
Introduction
Le changement démographique actuel dans les sociétés occidentales, impliquant à la fois une augmentation relative et absolue du nombre de personnes âgées, a suscité un intérêt scientifique croissant pour les questions gériatriques. Dans ce contexte, les concepts de vieillissement réussi deviennent de plus en plus importants. Plusieurs études ont indiqué que l’exercice physique pourrait jouer un rôle clé dans le vieillissement en santé et dans la prévention du déclin cognitif et des maladies neurodégénératives (Colcombe et al., 2006; Erickson et coll., 2011). Des revues récentes résumant des études épidémiologiques, transversales et interventionnelles soutiennent l’activité physique comme méthode propice pour induire la neuroplasticité à la fin de l’âge adulte (Gregory et al., 2012; Voelcker-Rehage et Niemann, 2013; Bamadis et al., 2014). Erickson et coll. (2012) ont conclu que des niveaux plus élevés d’aptitude cardiorespiratoire et d’activité physique aérobie sont associés à des volumes de matière grise plus importants dans les régions préfrontales et l’hippocampe. Cependant, non seulement la forme cardiovasculaire, mais aussi l’exercice de coordination (Niemann et al., 2014) et l’entraînement cognitif (Bamadis et al., 2014) induisent la plasticité de la matière grise et améliorent les fonctions cognitives chez les personnes âgées.
La recherche sur les animaux a suggéré qu’une combinaison d’exercice physique et d’enrichissement sensoriel a l’effet le plus fort sur la genèse de nouveaux neurones — principalement dans l’hippocampe — et que seule cette combinaison assure la survie durable des cellules nouveau-nées (Kempermann et al., 1997; van Praag et coll., 2005). Kattenstroth et coll. (2013) ont suggéré que « les activités de danse devraient être considérées comme un équivalent de conditions environnementales enrichies pour les humains car elles fournissent à un individu des exigences sensorielles, motrices et cognitives accrues. »Malgré cette déclaration encourageante, les études examinant les effets de l’entraînement en danse sur la structure ou la fonction du cerveau sont rares. Après une intervention dansante de 6 mois, Kattenstroth et al. (2010) ont signalé des améliorations significatives de la performance cognitive, tactile et motrice chez les personnes âgées participantes. Les résultats d’une étude prospective de 469 sujets âgés de plus de 75 ans sur une période médiane de suivi de 5,1 ans ont indiqué que la danse est associée à un risque nettement réduit de démence (Verghese et al., 2003). Cependant, Hüfner et coll. (2011) ont rapporté des volumes réduits dans plusieurs régions du cerveau, y compris la formation antérieure de l’hippocampe et des parties du cortex vestibulaire pariéto-insulaire chez les danseurs professionnels et les slackliners par rapport aux non-professionnels. Cette découverte peut indiquer que l’entraînement intensif et répétitif des mêmes habiletés motrices conduit dans un contexte de spécialisation à des volumes réduits de certaines régions du cerveau. Dans cette étude, nous concevons donc un programme d’entraînement spécial à la danse qui obligeait constamment les participants à apprendre de nouveaux schémas de mouvement (Müller et al., 2016). Pour évaluer les avantages spécifiques de cette intervention, nous avons comparé notre programme de danse nouvellement conçu à un groupe témoin actif plutôt qu’à un groupe témoin passif, qui participait à un programme de conditionnement physique classique dans lequel les participants effectuaient généralement des exercices physiques répétitifs tels que le vélo sur un ergomètre. De plus, parce que nous nous sommes intéressés à la dynamique temporelle des interventions, nous avons évalué les effets sur la structure et la fonction du cerveau après 6 et 18 mois d’entraînement. Ce faisant, nous avons cherché à évaluer s’il était bénéfique d’étendre les interventions, car certaines régions du cerveau peuvent nécessiter plus d’entraînement que d’autres, ou s’il existe une limite après laquelle plus d’entraînement devient préjudiciable. Enfin, à la recherche d’un mécanisme potentiel sous-jacent à la neuroplasticité, nous avons mesuré les taux de BDNF dans le sang périphérique. Plusieurs études ont suggéré que le BDNF favorise la différenciation de nouveaux neurones et synapses (Huang et Reichardt, 2001; Lessmann et Brigadski, 2009; Park et Poo, 2013; Edelmann et al., 2014). Il a donc été proposé que le BDNF soit un médiateur de la neuroplasticité chez l’adulte (Flöel et al., 2010).
Matériaux et méthodes
Participants et conception expérimentale
L’étude a été conçue comme une intervention contrôlée de 18 mois. L’étude a été approuvée par le comité d’éthique de l’Université Otto-von-Guericke de Magdebourg, et tous les sujets ont signé un écrit informé et ils n’ont pas reçu de paiement pour leur participation. Soixante-deux personnes âgées en bonne santé (63-80 ans) recrutées via des annonces dans les journaux locaux ont été sélectionnées pour l’étude. Les critères d’exclusion étaient la claustrophobie, les acouphènes, les implants métalliques, les tatouages, le diabète sucré, la dépression (Inventaire des dépressions de Beck, BDI-II > 13), les déficits cognitifs (Mini-Examen de l’état mental, MMSE < 27), les maladies neurologiques et l’exercice régulier (≥1 h / semaine). Sur la base de ces critères, 10 sujets ont été exclus. Les 52 participants restants ont ensuite été assignés au hasard à la danse ou au groupe sportif en utilisant le site Web www.randomization.com et en contrôlant l’âge, le statut MMSE et la forme physique. Les évaluations ont été effectuées au départ, après 6 et après 18 mois de formation (figure 1). Vingt-deux participants ont complété toute l’intervention et toutes les mesures. Le tableau 1 fournit des données démographiques détaillées pour ces participants. Aucune différence de groupe concernant les données démographiques n’a été constatée.
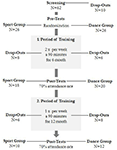
Figure 1. Organigramme du recrutement des participants.
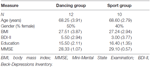
Tableau 1. Informations démographiques sur les participants au départ.
Interventions
Les interventions ont été séparées en deux périodes. Au cours de la première période, les sujets se sont entraînés deux fois par semaine en sessions de 90 minutes pendant 6 mois. Pour des raisons pratiques (disponibilité des participants et des formateurs), la deuxième période d’intervention de 12 mois comprenait une fréquence de formation réduite d’une fois par semaine. Les deux programmes d’intervention ont été exécutés dans un contexte de groupe avec de la musique pour contrôler les interactions psychosociales. La charge conditionnelle a été examinée en enregistrant les valeurs du pouls pendant les séances d’entraînement et en calculant la fréquence cardiaque d’entraînement individuelle selon Karvonen et al. (1957) avec un facteur 0,6 pour un entraînement intensif en endurance. Par conséquent, les deux programmes de formation étaient comparables en termes d’intensité, de durée et de fréquence. Les deux étaient supervisés par des instructeurs expérimentés.
Groupe de danse
Les participants du groupe de danse ont suivi un programme de formation nouvellement conçu dans lequel ils étaient constamment invités à apprendre de nouvelles séquences de mouvements. Ces chorégraphies nécessitaient la coordination de différentes parties du corps (c.-à-d. jambes, bras, tronc) dans l’espace sous différentes conditions de tension (tension physique, précision, pression de situation et de temps). Les sujets devaient apprendre les chorégraphies par cœur, imposant ainsi de hautes exigences à la mémoire. Le programme comprenait cinq genres différents (danse en ligne, danse jazz, rock « n » roll et danse carrée), qui ont été changés après chaque quatrième session. Au cours de l’intervention, les exigences de coordination et la pression du temps ont été accrues en introduisant des mouvements de danse et des chorégraphies plus complexes et en augmentant les battements par minute dans la musique.
Groupe sportif
Les participants au groupe sportif ont suivi un programme d’entraînement conventionnel en force-endurance avec des exercices principalement répétitifs et de faibles exigences en termes de coordination et de mémoire du corps entier. Chaque session comprenait des unités d’entraînement d’endurance, de force-endurance et de flexibilité de 20 minutes. L’entraînement d’endurance a été effectué sur des ergomètres à cycle. Dans l’unité force-endurance, des mouvements alternés (par exemple, boucles de biceps, squats, redressements assis) ont été effectués, mais des mouvements complexes du corps entier ont été évités pour maintenir les exigences de coordination faibles. L’unité de flexibilité consistait principalement en des exercices d’étirement.
Mesures des résultats
Condition cardiovasculaire
La condition cardiovasculaire a été évaluée par le Test de Capacité de travail Physique 130 (PWC 130). PWC 130 est la puissance de sortie (mesurée en watts) qu’un sujet est capable d’atteindre sur un ergomètre à cycle sous une fréquence cardiaque de 130 bpm. Pour les calculs, nous avons utilisé la fréquence cardiaque au repos et la capacité physique relative.
Tests neuropsychologiques
Une batterie étendue de tests neuropsychologiques a été réalisée sur les sujets. Aux fins de la présente étude, seuls les résultats d’un test de mémoire verbale à court et à long terme, le « VLMT » (une version allemande adaptée du « Test d’Apprentissage Verbal Auditif Rey »; Helmstaedter et Durwen, 1990) et d’une batterie de test d’attention (Test de Performance Attentionnelle (TAP); Zimmermann et Fimm, 2002) sont rapportés.
BDNF
Des échantillons de sang à jeun ont été prélevés le matin des évaluations neuropsychologiques. À partir des échantillons sanguins, les concentrations plasmatiques de BDNF ont été déterminées par des ELISAs sandwich (DuoSets BDNF; R& Systèmes D, Wiesbaden, Allemagne) comme décrit précédemment (Schega et al., 2016).
IRM
Des images IRM ont été acquises sur un MAGNÉTOM Verio Siemens de 3 Tesla (Syngo MR B17) à l’aide d’une bobine de tête à 32 canaux. Des séquences de MPRAGE pondérées en T1 à haute résolution ont été acquises à l’aide d’un protocole d’imagerie par écho à gradient rapide préparé par aimantation 3D (224 tranches sagittales, taille du voxel: 0,8 × 0,8 × 0,8 mm3, TR: 2500 ms, TE: 3,47 ms, TI: 1100 ms, angle de retournement: 7°). Les images MR ont été analysées à l’aide de la morphométrie basée sur le voxel (VBM) implémentée dans SPM 12 (Welcome Department of Cognitive Neurology, Londres, Royaume-Uni). La VBM est une technique impartiale du cerveau entier pour l’analyse du volume régional de matière grise et des changements tissulaires (Ashburner et Friston, 2000).
Le prétraitement impliquait la segmentation de la matière grise, la création de modèles via DARTEL, la normalisation spatiale dans l’espace normalisé de l’Institut neurologique de Montréal (INM) et le lissage avec un noyau gaussien de 8 mm de largeur totale à moitié maximum (FWHM).
Pour analyser la différence dans les changements de volume de matière grise entre les groupes, un plan factoriel complet avec le groupe de facteurs (danse, sport) et le temps (0, 6 et 18 mois) a été appliqué. Dans le cas d’interactions significatives de groupe × temps, des tests t post-hoc entre des paires consécutives de points temporels (0 vs 6; 0 vs 18; 6 vs. 18 mois) ont été calculés séparément pour chaque groupe. Un seuil de p< 0,001 (non corrigé) a été appliqué pour toutes les analyses.
Résultats
La présentation des résultats est structurée comme suit. Nous avons d’abord cherché un effet d’intervention général (facteur temps), puis recherché des effets différentiels des deux interventions (groupe d’interaction × temps); enfin, une analyse plus détaillée de la dynamique temporelle a été réalisée via des comparaisons post-hoc par paires.
Fitness cardiovasculaire
La fitness cardiovasculaire mesurée par le PWC 130 (Tableau 2, capacité physique relative) ne différait pas d’un groupe à l’autre au départ. De plus, le score PWC 130 n’a pas augmenté de manière significative pendant la durée de l’une ou l’autre intervention.

Tableau 2. Moyens et (SD) pour la forme physique, le fonctionnement cognitif, les niveaux plasmatiques de BDNF et le volume total de matière grise au sein des groupes d’entraînement au cours de l’intervention.
Tests neuropsychologiques
Un effet principal significatif du temps a été observé pour la mémoire verbale à court terme (rappel précoce VLMT; F(2,19) = 6,438, p = 0,004, η2 = 0,253), rappel libre verbal à long terme (rappel tardif VLMT; F (2,19) = 3,387, p = 0,049, η2 = 0,244), rappel long verbal – reconnaissance du terme (reconnaissance VLMT; F(2,19) = 5,352, p = 0,009, η2 = 0,220) et temps de réaction de l’attention (flexibilité du sous-test; F(2,19) = 19,156, p < 0,001, η2 = 0,489). Aucune interaction significative de temps × de groupe n’est apparue.
Les comparaisons post-hoc par paires ont montré des améliorations significatives de la valeur de référence à 18 mois et de 6 à 18 mois dans les trois sous-catégories de VLMT dans les deux groupes. En ce qui concerne les temps de réaction au TAP, une amélioration significative a été observée dans la comparaison de la valeur de référence avec les données sur 18 mois dans les deux groupes.
BDNF
Les taux plasmatiques de BDNF ont été analysés dans des échantillons de sang avant le début de l’entraînement ainsi que 6 mois et 18 mois après le début de l’entraînement. Les concentrations plasmatiques absolues de BDNF sont résumées dans le tableau 2. Les changements intra-individuels du niveau de BDNF ont révélé une augmentation significative du groupe de danse, alors que le niveau intra-individuel de BDNF est resté constant après 6 mois d’entraînement dans le groupe sportif. Aucun autre changement dans le BDNF n’est survenu après 18 mois dans aucun groupe (figure 2).
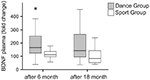
Figure 2. Changements intra-individuels des taux plasmatiques de BDNF après intervention. Les taux plasmatiques de BDNF ont été analysés dans des échantillons de sang de participants effectuant un programme d’entraînement en danse ou un programme d’entraînement sportif avant le début de l’entraînement, après 6 mois d’entraînement et après 18 mois d’entraînement. L’augmentation relative des niveaux de BDNF a été quantifiée. Les niveaux de BDNF ont augmenté de manière significative dans le groupe de danse après 6 mois d’entraînement (test U de Mann-Whitney, p < 0,004) et ont diminué presque jusqu’à la valeur de base de la valeur de prétraitement après 18 mois d’entraînement en danse. Il n’y a pas eu de changement sur toute la durée du parcours dans le groupe sportif (test de Friedman, p = 0,319), alors qu’un changement significatif sur toute la durée du parcours a été observé dans le groupe de danse (test de Friedman, p = 0,028). Placettes : minimum, 25e percentile, médiane, 75e percentile. * p ≤ 0,05.
IRM
Une interaction significative de groupe ×temps a été observée dans le gyrus précentral gauche et le parahippocampe droit. Les tests t post hoc entre les données de base et les données de 6 mois ont montré une augmentation significative du volume de matière grise dans le gyrus précentral gauche de seulement les danseurs. En comparant les données de base et les données sur 18 mois en plus du gyrus précentral, les danseurs ont présenté une augmentation significative du volume de matière grise dans le gyrus parahippocampique droit, qui était également le seul changement significatif dans l’intervalle de 6 à 18 mois. Ainsi, l’augmentation du volume du gyrus précentral est apparue après 6 mois et est restée stable pendant l’intervalle d’entraînement de danse restant, alors que le changement du gyrus parahippocampique s’est produit uniquement pendant l’intervalle d’entraînement ultérieur (figure 3).
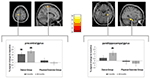
Figure 3. Analyse du temps par interaction de groupe, test pour des changements de volume plus importants dans la danse par rapport au groupe sportif. Une augmentation significative de la matière grise a été observée dans le gyrus précentral (coordonnées de l’Institut neurologique de Montréal (INM) : x =-16; y =-18; z = 77) et dans le gyrus parahippocampique (coordonnées de l’INM : x =34; y =-26; z =-20). Les diagrammes de boîte montrent les changements relatifs de matière grise dans le voxel de crête. * p ≤ 0,05.
Discussion
Dans cette étude, nous avons comparé les effets de la participation à un programme de danse ou à un programme de sport de forme physique conventionnel sur le fonctionnement et le volume du cerveau chez les personnes âgées en bonne santé. Le programme de danse était une intervention nouvellement conçue qui nécessitait d’apprendre constamment de nouvelles chorégraphies de danse. Le programme de sport conventionnel était principalement axé sur des exercices moteurs répétitifs. Comme principale constatation, nous avons observé qu’après 6 mois d’entraînement, les volumes dans le gyrus précentral gauche des danseurs avaient augmenté plus que ceux du groupe sportif. Après encore 12 mois d’entraînement, une augmentation de volume supplémentaire a été observée dans le gyrus parahippocampique droit des danseurs. Les niveaux de BDNF ont augmenté au cours des 6 premiers mois d’entraînement en danse et sont revenus aux valeurs de prétraitement après 18 mois. Dans le groupe de sport conventionnel, une augmentation similaire du BDNF n’était pas évidente. Étant donné que les niveaux de forme cardiovasculaire au cours des interventions sont restés constants dans les deux groupes, les effets observés ne pouvaient pas être attribués à des améliorations de la forme physique, mais semblaient plutôt être liés aux caractéristiques spécifiques du programme de danse. Ces caractéristiques comprenaient l’exigence d’apprendre constamment de nouvelles chorégraphies (c’est-à-dire la mémoire), d’intégrer des informations multisensorielles, de coordonner tout le corps et de naviguer dans l’espace.
Changements cérébraux
Le gyrus précentral est essentiel au contrôle des fonctions motrices volontaires. L’augmentation du volume de matière grise dans le gyrus précentral dans le groupe de danse peut donc être basée sur les schémas de mouvement complexes et en constante évolution que les danseurs devaient effectuer. Ces mouvements nécessitaient la coordination simultanée de plusieurs parties du corps dans des directions différentes et l’adaptation aux rythmes variables de la musique (polycentrique et polyrythmique). En réfléchissant à ces exigences complexes de coordination, Brown et coll. (2006) ont rapporté des activations induites par la danse dans le putamen, le cortex moteur primaire et la zone motrice supplémentaire (SMA), comme le montre la TEP. D’autres études ont indiqué une association entre les exigences de coordination (p. ex., équilibrage, jonglage) et la neuroplasticité dans la région précentrale (Boyke et al., 2008; Taubert et coll., 2010). Par conséquent, l’augmentation du volume lié à la danse dans ce domaine était conforme aux attentes fondées sur la littérature.
Le gyrus parahippocampique fait partie de l’arc externe du système limbique et joue un rôle important dans la mémoire de travail et la récupération de la mémoire épisodique (Pantel et al., 2003). Selon Bliss et Lomo (1973), le gyrus parahippocampique constitue l’interface entre la mémoire et la conscience expérientielle du présent, car il est interconnecté par le tractus perforant à la fois aux régions du lobe frontal, associées à la mémoire de travail, et à l’hippocampe, la structure centrale du codage de la mémoire épisodique et de la navigation spatiale. De nombreuses études VBM ont rapporté une perte de volume liée à l’âge dans les régions parahippocampiques (Tisserand et al., 2002). En outre, Echávarri et coll. (2011) ont suggéré que l’atrophie parahippocampique est un biomarqueur précoce de la MA. Le constat que s’engager dans un programme de danse sur une plus longue période peut induire des processus neuroplastiques dans cette région cruciale de la mémoire, donc particulièrement encourageant en termes de développement de stratégies de prévention.
Dynamique temporelle de la Plasticité cérébrale de la Matière grise
Le volume observé augmente dans les deux régions cérébrales développées à des moments différents. La danse a entraîné une augmentation du volume des zones motrices après 6 mois (voir également Rehfeld et al., 2016), qui est restée stable au cours des 12 mois suivants. L’augmentation de volume associée à la danse dans le gyrus parahippocampique est apparue plus tard et n’a été observée que dans les données de 18 mois. Les différentes dynamiques temporelles dans l’évolution des deux régions cérébrales peuvent être liées à des différences dans les mécanismes cellulaires sous-jacents. La recherche sur les animaux a suggéré que l’angiogenèse et la formation de nouvelles dendrites (Thomas et al., 2012) se produisent rapidement, alors que les changements dans le neuropile se produisent beaucoup plus lentement (Black et al., 1990). Chez l’homme, des augmentations rapides du volume de matière grise dans les régions préfrontales ont été observées après seulement 2 semaines d’apprentissage moteur chez les jeunes adultes (Taubert et al., 2010). Pour induire une neuroplasticité dans l’hippocampe, des périodes d’entraînement plus longues ont été recommandées (Erickson et al., 2012; Niemann et coll., 2014).
Fait intéressant, les augmentations de volume susmentionnées de notre étude contrastent avec les résultats de Hänggi et al. (2010), qui a révélé que les danseurs de ballet professionnels ont diminué les volumes de matière grise dans le cortex prémoteur gauche, la SMA, le Putamen et le gyrus frontal supérieur et les résultats de Hüfner et al. (2011), qui rapportent des volumes réduits dans plusieurs régions du cerveau, y compris la formation antérieure de l’hippocampe chez les danseurs professionnels et les slackliners par rapport aux non-professionnels. Cependant, ces études étaient des études observationnelles transversales qui comparaient le cerveau des danseurs professionnels à celui des non-professionnels. De plus, notre groupe de danse était composé de novices âgés. Il a été démontré que l’apprentissage d’une nouvelle compétence conduit d’abord au recrutement de ressources neuronales supplémentaires. Plus tard, lorsque la compétence devient plus automatique, moins de ressources neuronales sont nécessaires, ce qui peut entraîner une diminution du volume chez ceux qui ont une expérience à long terme. Sur la base de ces conditions initiales (novices, adultes plus âgés) et de notre programme d’entraînement à la danse spécialement conçu, qui nécessitait un apprentissage constant des schémas de mouvement, il est possible que les effets de la spécialisation induisent une diminution du volume, comme le rapportent Hänggi et al. (2010) ne sont pas observées.
On pense généralement que l’entraînement moteur induit initialement une augmentation du volume cérébral. Cependant, un entraînement prolongé conduit à une automatisation, qui peut avoir les effets opposés sur le volume cortical, car un contrôle cortical moins important est nécessaire une fois que les compétences motrices ont été pleinement établies (Taube, 2008). Notre programme d’entraînement en danse a donc été spécialement conçu pour éviter une telle automatisation, ce qui peut expliquer pourquoi, au moins dans les 18 mois, aucune diminution du volume cortical n’a été observée dans notre étude.
Les fonctions cognitives ont également montré un développement non linéaire, la mémoire verbale n’augmentant que pendant la deuxième période d’entraînement. En ce qui concerne la performance de l’attention, des améliorations significatives étaient observables après seulement 6 mois dans les deux groupes (Rehfeld et al., 2016). Ces résultats confirment les rapports antérieurs concernant les effets bénéfiques des interventions physiques sur les tests neuropsychologiques (Bamadis et al., 2014). Cependant, dans les données sur les capacités cognitives, contrairement aux données sur le cerveau, aucune différence de groupe n’est apparue. D’autres ont signalé des effets supérieurs de l’entraînement cognitif et physique combiné par opposition à des interventions uniques (Oswald et al., 2006). Nous étendrons nos interventions pour tester si des différences de cognition de groupe pourraient apparaître à des moments encore plus tardifs.
Mécanismes cellulaires et moléculaires sous-jacents de la plasticité de la Matière grise
Bien que la MBV soit une modalité d’imagerie qui révèle des changements de volume dans le cerveau, cette technique ne permet pas de conclusions causales concernant les processus neurophysiologiques sous-jacents. La neurogenèse, la synaptogenèse et l’angiogenèse ne sont que quelques-uns des mécanismes qui ont été suggérés comme étant à la base des changements de volume cérébral (Zatorre et al., 2012). En tant que médiateurs des effets de la forme cardiovasculaire sur le cerveau, des facteurs de croissance tels que le BDNF, le facteur de croissance analogue à l’insuline (IGF) et le facteur de croissance nerveuse (NGF) sont à l’étude (Kirk-Sanchez et McGough, 2014). Cependant, dans notre étude, contrairement aux précédentes (Erickson et al., 2011; Maass et coll., 2015), aucune différence de condition physique cardiovasculaire n’était présente entre les groupes et les niveaux de condition physique n’ont pas changé au cours des interventions. Cette dernière observation était probablement liée à notre contrôle de la fréquence cardiaque individuelle, que nous voulions maintenir dans la zone aérobie. Cependant, les changements de BDNF ont également été associés à l’activité physique, à l’interaction sociale et au stress positif (Mattson, 2008), et toutes les études n’ont pas observé une augmentation de BDNF après un entraînement cardiovasculaire (Vital et al., 2014). Enfin, la recherche sur les animaux a suggéré que la coordination, mais pas l’entraînement en endurance, induit une synaptogenèse et des changements gliaux (Black et al., 1990). Ensemble, les facteurs supplémentaires nommés qui stimulent la sécrétion de BDNF peuvent avoir été plus cruciaux pendant la danse que pendant les activités de fitness, expliquant ainsi pourquoi seuls les danseurs ont montré une augmentation de BDNF au cours des 6 premiers mois. L’observation selon laquelle les niveaux de BDNF sont revenus au niveau de référence au cours des 12 mois suivants, tandis que des augmentations de volume ont été observées simultanément dans le gyrus parahippocampique, indique cependant qu’il doit y avoir d’autres facteurs impliqués dans la plasticité cérébrale de l’adulte que ceux représentés par les niveaux de BDNF dans le sang périphérique.
En ce qui concerne les mécanismes neurobiologiques de la plasticité induite par l’exercice, il existe également un concept de réserve cérébrale (Satz et al., 2011) devrait être envisagée. Le concept de réserve cérébrale décrit les différences individuelles dans une neuroplasticité adaptative de base accrue, qui fournissent une plus grande capacité dynamique de remodelage des circuits corticaux en différents facteurs de stress (Barulli et Stern, 2013; Freret et al., 2015).
Perspectives
Les résultats de notre étude suggèrent qu’une intervention de danse à long terme pourrait être supérieure à un exercice physique répétitif pour induire une neuroplasticité dans le cerveau humain vieillissant. Nous supposons que cet avantage est lié à la nature multimodale de la danse, qui combine des défis physiques, cognitifs et de coordination. À notre connaissance, il s’agit de la première étude longitudinale randomisée à recommander des programmes de danse comme moyen de prévenir la matière grise et le déclin cognitif chez les personnes âgées. D’autres recherches sont nécessaires pour clarifier plus en détail la dynamique temporelle et les mécanismes neurobiologiques sous-jacents de la neuroplasticité induite par la danse et si cette intervention a vraiment le potentiel de réduire le risque de maladies neurodégénératives telles que la maladie d’Alzheimer.
Contributions des auteurs
PM a conçu et réalisé la recherche, analysé les données, écrit l’article. KR a conçu et réalisé la recherche. MS a analysé les données. AH et VL ont conçu la recherche. MD: collecte de données, révision d’articles. TB et JK ont conçu la recherche, analysé les données. NGM a conçu la recherche, a écrit l’article.
Déclaration sur les conflits d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de relations commerciales ou financières pouvant être interprétées comme un conflit d’intérêts potentiel.
Ashburner, J., et Friston, K. J. (2000). Morphométrie basée sur le voxel – les méthodes. Neuroimage 11, 805-821. doi: 10.1006 / nimg.2000.0582
Résumé PubMed | CrossRef Texte intégral / Google Scholar
Bamadis, P. D., Vivas, A. B., Styliadis, C., Frantzidis, M., Klados, M., Schlee, W., et al. (2014). Une revue des interventions physiques et cognitives dans le vieillissement. Neurosci. Biocomportement. Rév.44, 206-220. doi: 10.1016/ j.neubiorev.2014.03.019
Résumé PubMed | Texte intégral CrossRef /Google Scholar
Barulli, D., et Stern, Y. (2013). Efficacité, capacité, compensation, maintenance, plasticité: concepts émergents dans la réserve cognitive. Tendances Cogn. Sci. 17, 502–509. doi: 10.1016/ j. tics.2013.08.012
Résumé publié par PubMed | Texte intégral croisé / Google Scholar
Black, J. E., Isaacs, K. R., Anderson, B. J., Alcantara, A.A., et Greenough, W. T. (1990). L’apprentissage provoque la synaptogenèse, tandis que l’activité motrice provoque l’angiogenèse, dans le cortex cérébelleux des rats adultes. Proc. Natl. Acad. Sci. U S A 87, 5568-5572. doi: 10.1073/pnas.87.14.5568
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Bliss, T. V., et Lomo, T. (1973). Potentialisation de longue durée de la transmission synaptique dans le denté du lapin anéastétisé suite à la stimulation de la voie perforante. J. Physiol. 232, 331–356. doi: 10.1113/jphysiol.1973.sp010273
Résumé publié | Texte intégral croisé / Google Scholar
Boyke, J., Driemeyer, J., Gasser, C., Büchel, C. et May, A. (2008). Changements de structure cérébrale induits par l’entraînement chez les personnes âgées. J. Neurosci. 28, 7031–7035. doi: 10.1523/JNEUROSCI.0742-08.2008
Résumé PubMed | Texte intégral CrossRef / Google Scholar
Brown, S., Martinez, M. et Parsons, M. (2006). La base neuronale de la danse humaine. Céreb. Cortex 16, 1157-1167. doi: 10.1093/cercor/bhj057
Résumé publié | Texte intégral croisé / Google Scholar
Colcombe, S. J., Erickson, K. I., Scalf, P. E., Kim, J. S., Prakash, R., McAuley, E., et al. (2006). L’entraînement par exercices aérobiques augmente le volume cérébral chez les humains vieillissants. J. Gerontol. A. Biol. Sci. Med. Sci. 61, 1166–1170. doi: 10.1093/gérone/61.11.1166
Résumé publié | Texte intégral croisé / Google Scholar
Echávarri, C., Aalten, P., Uyling, H. B. M., Jacobs, H. I. L., Visser, P. J., Gronenschild, M., et al. (2011). Atrophie du gyrus parahippocampique en tant que biomarqueur précoce de la maladie d’Alzheimer. Structure cérébrale. Fonction. 215, 265–271. doi: 10.1007/s00429-010-0283-8
Résumé publié par PubMed / Texte intégral croisé / Google Scholar
Edelmann, E., Lessmann, V. et Brigadski, T. (2014). Torsions pré et postsynaptiques dans la sécrétion de BDNF et l’action dans la plasticité synaptique. Neuropharmacologie 76, 610-627. doi: 10.1016/j. neuropharm.2013.05.043
Résumé PubMed | Texte intégral croisé / Google Scholar
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., et al. (2011). L’entraînement physique augmente la taille de l’hippocampe et améliore la mémoire. Proc. Natl. Acad. Sci. U S A 108, 3017-3022. doi: 10.1073/pnas.1015950108
Résumé PubMed | Texte intégral CrossRef /Google Scholar
Erickson, K. I., Weinstein, A. M. et Lopez, O.L. (2012). Activité physique, plasticité cérébrale et maladie d’Alzheimer. Arch. Med. Rés. 43, 615 à 621. doi: 10.1016 /j. arcmé.2012.09.008
Résumé PubMed | CrossRef Texte intégral / Google Scholar
Flöel, A., Ruscheweyh, R., Krüger, K., Willemer, C., Winter, B., Völker, K., et al. (2010). Activité physique et fonctions de la mémoire: les neurotrophines et le volume de matière grise cérébrale sont-ils le chaînon manquant? Neuroimage 49, 2756-2763. doi: 10.1016/j. neuroimage.2009.10.043
Résumé PubMed / Texte intégral croisé / Google Scholar
Freret, T., Gaudreau, P., Schumann-Bard, P., Billard, J. M., et Popa-Wagner, A. (2015). Mécanismes sous-jacents à l’effet neuroprotecteur de la réserve cérébrale contre la dépression en fin de vie. J. Neural. Transm. (Vienne) 122, S55-S61. doi: 10.1007/s00702-013-1154-2
Résumé PubMed | Texte intégral CrossRef / Google Scholar
Gregory, S. M., Parker, B., et Thompson, P.D. (2012). Activité physique, fonction cognitive et santé du cerveau: quel est le rôle de l’entraînement physique dans la prévention de la démence? Cerveau Sci. 2, 684–708. doi: 10.3390/brainsci2040684
Résumé publié / Texte intégral croisé /Google Scholar
Hänggi, J., Koeneke, S., Bezzola, L., et Jäncke, L. (2010). Neuroplasticité structurelle dans le réseau sensorimoteur de danseuses de ballet professionnelles. Hum. Mapp cerveau. 31, 1196–1206. doi: 10.1002/hbm.20928
Résumé PubMed / Texte intégral croisé / Google Scholar
Helmstaedter, C., et Durwen, H. F. (1990). Le test d’apprentissage verbal et de rétention. Un outil utile et différencié pour évaluer les performances de la mémoire verbale. Suisse. Arch. Neurol. Psychiatre. (1985) 141, 21–30.
Résumé PubMed /Google Scholar
Huang, E. J., et Reichardt, L. F. (2001). Neurotrophines: rôles dans le développement et la fonction neuronaux. Annu. Rév. Neurosci. 24, 677–736. doi: 10.1146/annurev.neuro.24.1.677
Résumé publié par PubMed / CrossRef Texte intégral / Google Scholar
Hüfner, K., Binetti, C., Hamilton, D. A., Stephan, T., Flanagin, V. L., Linn, J., et al. (2011). Plasticité structurelle et fonctionnelle de la formation de l’hippocampe chez les danseurs professionnels et les slackliners. Hippocampe 21, 855-865. doi: 10.1002/ hipo.20801
Résumé publié par PubMed | Texte intégral croisé / Google Scholar
Karvonen, M. J., Kentala, E. et Mustala, O. (1957). Les effets de l’entraînement sur la fréquence cardiaque: une étude longitudinale. Ann. Med. Exp. Biol. Fenn. 35, 307–315.
Résumé PubMed /Google Scholar
Kattenstroth, J. C., Kalisch, T., Holt, S., Tegethoff, M., et Dinse, H. R. (2013). Six mois d’intervention en danse améliorent les performances posturales, sensorimotrices et cognitives chez les personnes âgées sans affecter les fonctions cardio-respiratoires. Devant. Neurosci vieillissants. 5:5. doi: 10.3389/ fnagi.2013.00005
Résumé PubMed | Texte intégral croisé / Google Scholar
Kattenstroth, J. C., Kolankowska, I., Kalisch, T., et Dinse, HR (2010). Performances sensorielles, motrices et cognitives supérieures chez les personnes âgées avec des activités de danse pluriannuelles. Devant. Neurosci vieillissants. 2:31. doi: 10.3389/ fnagi.2010.00031
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Kempermann, G., Kuhn, H. G. et Gage, F. H. (1997). Plus de neurones hippocampiques chez des souris adultes vivant dans un environnement enrichi. Nature. 386, 493–495. doi: 10.1038/ 386493a0
Résumé PubMed | Texte intégral croisé | Google Scholar
Kirk-Sanchez, NJ, et McGough, El (2014). Exercice physique et performance cognitive chez les personnes âgées: perspectives actuelles. Clin. Interv. Vieillissement 9, 51-62. doi: 10.2147 / CIA.S39506
Résumé PubMed / Texte intégral croisé / Google Scholar
Lessmann, V., et Brigadski, T. (2009). Mécanismes, emplacements et cinétiques de la sécrétion synaptique de BDNF: une mise à jour. Neurosci. Rés. 65, 11 à 22. doi: 10.1016 / j.neures.2009.06.004
Résumé PubMed / CrossRef Texte intégral / Google Scholar
Maass, A., Düzel, S., Goerke, M., Becke, A., Sobieray, U., Neumann, K., et al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M., Hökelmann, A., Kaufmann, J., et al. (2016). Effets une danse et un entraînement sportif de santé sur la substance grise du cerveau des personnes âgées en bonne santé. Sportwiss. 46, 213-222. doi: 10.1007 / s12662-016-0411-6
CrossRef Full Text/Google Scholar
Niemann, C., Godde, B., And Voelcker-Rehage, C. (2014). Not only cardiovascular, but also coordinative exercise increases hippocampal volume in older adults. Avant. Vieillissement Neurosci. 6: 170. doi: 10.3389/fnagtp2014.00170
PubMed Abstract | CrossRef Full Text/Google Scholar
Oswald, W. D., Gunzelmann, T., Rupprecht, R., et Hagen, B. (2006). Effets différentiels de l’entraînement cognitif et physique combiné avec des adultes plus âgés: l’étude SimA dans une perspective de 5 ans. EUR. J. Vieillissement 3, 179-192. doi: 10.1007/s10433-006-0035- z
CrossRef Texte intégral /Google Scholar
Pantel, J., Kratz, B., Essig, M. et Schröde, J. (2003). Déficits volumiques parahippocampiques chez les sujets présentant un déclin cognitif associé au vieillissement. Être. J. Psychiatrie 160, 379-382. doi: 10.1176/appi.l’ajp.160.2.379
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Park, H., et Caca, M. M. (2013). Régulation des neurotrophines du développement et de la fonction des circuits neuronaux. NAT. Rév. Neurosci. 14, 7–23. doi: 10.1038/nrn3379
Résumé publié|Texte intégral croisé/Google Scholar
Rehfeld, K., Hökelmann, A., Kaufmann, J. et Müller, N. G. (2016). « Effects of a dance vs. a fitness training on brain plasticity, balance performance and attention in healthy seniores: a new approach with SPM 12 for pairwise longitudinal group comparison », in Proceeding – International Conference of Sports and Neuroscience (Magdebourg), 127-138.
Satz, P., Cole, M. A., Hardy, D. J. et Rassovsky, Y. (2011). Réserve cérébrale et cognitive : médiateur(s) et validité de la construction, une critique. J. Clin. Exp. Neuropsychol. 33, 121–130. doi: 10.1080 / 13803395.2010.493151
Résumé publié / Texte intégral croisé / Google Schega, L., Peter, B., Brigadski, T., Leßmann, V., Isermann, B., Hamacher, D., et al. (2016). Effet de l’hypoxie normobare intermittente sur la capacité aérobie et la fonction cognitive chez les personnes âgées. J. Sci. Med. Sport 19, 941-945. doi: 10.1016/ j.jsams.2016.02.012
Résumé PubMed / Texte intégral CrossRef /Google Scholar
Taube, W. (2008). Adaptations neurophysiologiques en réponse à l’entraînement à l’équilibre. Dtsch. Z. Sportmed. 63, 273–277. doi: 10.5960/ dzsm.2012.030
CrossRef Texte intégral / Google Scholar
Taubert, M., Draganski, B., Anwander, A., Müller, K., Horstmann, A., Villringer, A., et al. (2010). Propriétés dynamiques de la structure du cerveau humain: apprentissage des changements liés aux zones corticales et des connexions de fibres associées. J. Neurosci. 30, 11670–11677. doi: 10.1523/JNEUROSCI.2567-10.2010
Résumé PubMed | Texte intégral CrossRef / Google Scholar
Thomas, A. G., Dennis, A., Bandettini, P.A., et Johansen-Berg, H. (2012). Les effets de l’activité aérobie sur la structure du cerveau. Devant. Psychol. 3:86. doi: 10.3389 / fpsyg.2012.00086
Résumé publié par PubMed | Texte intégral croisé / Google Scholar
Tisserand, D. J., Pruessner, J. C., Sanz Arigita, E. J., van Boxtel, M. P., Evans, A. C., Jolles, J., et al. (2002). Les volumes corticaux frontaux régionaux diminuent de manière différentielle au cours du vieillissement: une étude IRM pour comparer les approches volumétriques et la morphométrie basée sur le voxel. Neuroimage 17, 657-669. doi: 10.1016/s1053-8119 (02) 91173-0
Résumé PubMed / Texte intégral croisé / Google Scholar
van Praag, H., Shubert, T., Zhao, C. et Gage, F. H. (2005). L’exercice améliore l’apprentissage et la neurogenèse de l’hippocampe chez les souris âgées. J. Neurosci. 25, 8680–8685. doi: 10.1523/JNEUROSCI.1731-05.2005
Résumé publié par PubMed / CrossRef Texte intégral / Google Scholar
Verghese, J., Lipton, R. B., Katz, M. J., Hall, C. B., Dervy, C. A., Kuslansky, G., et al. (2003). Les activités de loisirs et le risque de démence chez les personnes âgées. N. Engl. J. Med. 348, 2508–2516. doi: 10.1056/NEJMoa022252
Résumé publié / Texte intégral croisé / Google Scholar
Vital, T. M., Stein, A. M., de Melo Coelho, F. G., Arantes, F. J., Teodorov, E., Santos-Galduróz, R. F., et al. (2014). L’exercice physique et le facteur de croissance endothélial vasculaire (VEGF) chez les personnes âgées: une revue systématique. Arch. Gerontol. Gériatr. 59, 234–239. doi: 10.1016 / j. archger.2014.04.011
Résumé PubMed | CrossRef Texte intégral /Google Scholar
Voelcker-Rehage, C., et Niemann, C. (2013). Changements cérébraux structurels et fonctionnels liés à différents types d’activité physique tout au long de la vie. Neurosci. Biocomportement. Rév. 37, 2268-2295. doi: 10.1016/ j.neubiorev.2013.01.028
Résumé PubMed | Texte intégral CrossRef /Google Scholar
Zatorre, R. J., Fields, R. D., et Johansen-Berg, H. (2012). Plasticité en gris et blanc: changements de neuroimagerie dans la structure du cerveau pendant l’apprentissage. NAT. Neurosci. 15, 528–536. doi: 10.1038/nn.3045
PubMed Abstract | CrossRef Full Text/Google Scholar
Zimmermann, P., and Fimm, B. (2002). Batterie de test pour le contrôle de L’attention (TAP), Version 1.7, partie Manuelle1. Würselen: Psytest.