
Introduksjon
Den nåværende demografiske endringen I Vestlige samfunn, som involverer både en relativ og en absolutt økning i antall eldre mennesker, har skapt økende vitenskapelig interesse for geriatriske problemer. I denne sammenheng blir begreper om vellykket aldring stadig viktigere. Flere studier har indikert at fysisk trening kan spille en nøkkelrolle i sunn aldring og i forebygging av kognitiv tilbakegang og nevrodegenerative sykdommer (Colcombe et al., 2006; Erickson et al., 2011). Nylige anmeldelser som oppsummerer epidemiologiske, tverrsnittsstudier og intervensjonsstudier, støtter fysisk aktivitet som en gunstig metode for å indusere nevroplastisitet i sen voksen alder (Gregory et al., 2012; Voelcker-Rehage og Niemann, 2013; Bamadis et al., 2014). Erickson et al. (2012) har konkludert med at høyere kardiorespiratorisk kondisjon og aerob fysisk aktivitetsnivå er forbundet med større gråstoffvolumer i prefrontale regioner og hippocampus. Men ikke bare kardiovaskulær kondisjon, men også koordinativ trening (Niemann et al., 2014) og kognitiv trening (Bamadis et al., 2014) har vist seg å indusere grå materie plastisitet og forbedre kognitive funksjoner hos eldre voksne.Dyreforskning har antydet at en kombinasjon av fysisk trening med sensorisk berikelse har den sterkeste effekten på genesis av nye nevroner-hovedsakelig i hippocampus-og at bare denne kombinasjonen sikrer varig overlevelse av nyfødte celler (Kempermann et al., 1997; van Praag et al., 2005). Kattenstroth et al. (2013) har antydet at «dansaktiviteter bør betraktes som en ekvivalent av berikede miljøforhold for mennesker, siden de gir et individ økte sensoriske, motoriske og kognitive krav.»Til tross for denne oppmuntrende uttalelsen er studier som undersøker effekten av dansetrening på hjernestruktur eller funksjon, knappe. Etter en 6-måneders danseintervensjon, Kattenstroth et al. (2010) har rapportert betydelige forbedringer i kognitiv, taktil og motorisk ytelse hos deltakende eldre. Resultatene av en prospektiv studie av 469 personer eldre enn 75 år over en median oppfølgingsperiode på 5,1 år har indikert at dans er forbundet med en markert redusert risiko for demens (Verghese et al., 2003). Imidlertid Er Hü et al. (2011) har rapportert reduserte volumer i flere hjernegrupper, inkludert den fremre hippocampale formasjonen og deler av parieto-insulære vestibulære cortex hos profesjonelle dansere og slackliner sammenlignet med ikke-profesjonelle. Dette funnet kan tyde på at intensiv og repeterende trening av de samme motoriske ferdighetene fører i sammenheng med spesialisering til reduserte volumer av noen hjernegrupper. I denne studien utformer vi derfor et spesielt dansetreningsprogram som hele tiden krevde at deltakerne lærte nye bevegelsesmønstre (Mü et al., 2016). For å vurdere de spesifikke fordelene med denne intervensjonen, sammenlignet vi vårt nyutviklede danseprogram med en aktiv snarere enn en passiv kontrollgruppe, som deltok i et konvensjonelt helsesport-treningsprogram der deltakerne vanligvis utførte repeterende fysiske øvelser som sykling på et ergometer. Videre, fordi vi var interessert i den tidsmessige dynamikken i intervensjonene, vurderte vi effektene på hjernestruktur og funksjon etter 6 og 18 måneders trening. Ved å gjøre det, søkte vi å vurdere om det er gunstig å utvide intervensjoner, fordi noen hjernegrupper kan kreve mer trening enn andre, eller om det er en grense etter hvilken mer trening blir skadelig. Til slutt, på jakt etter en potensiell mekanisme som ligger til grunn for neuroplasticitet, målte VI bdnf-nivåene i perifert blod. Flere studier har antydet AT BDNF fremmer differensiering av nye nevroner og synapser (Huang Og Reichardt, 2001; Lessmann Og Brigadski, 2009; Park and Poo, 2013; Edelmann et al., 2014). BDNF, derfor, har blitt foreslått å være en mellommann for voksen neuroplasticity (Fl@el et al., 2010).
Materialer og Metoder
Deltakere og Eksperimentell Design
studien ble utformet som en 18-måneders kontrollert intervensjon. Studien ble godkjent av Etikkutvalget Ved Otto-von-Guericke University, Magdeburg, og alle fag signerte en skriftlig informert og de mottok ikke betaling for deres deltakelse. Sixty-to friske eldre personer (63-80 år) rekruttert via kunngjøringer i lokale aviser ble screenet for studien. Eksklusjonskriteriene var klaustrofobi, tinnitus, metallimplantater, tatoveringer, diabetes mellitus, depresjon (Beck-Depression Inventory, BDI-II > 13), kognitive underskudd (Mini-Mental Tilstandsundersøkelse, MMSE < 27), nevrologiske sykdommer og regelmessig trening (≥1 time/uke). På grunnlag av disse kriteriene ble 10 fag ekskludert. De resterende 52 deltakerne ble deretter tilfeldig tildelt enten dans eller sport gruppe ved hjelp av nettstedet www.randomization.com og kontrollere for alder, MMSE status og fysisk form. Vurderinger ble utført ved baseline, etter 6 og etter 18 måneders trening (Figur 1). Tjueto deltakere fullførte hele intervensjonen og alle målinger. Tabell 1 gir detaljerte demografiske data for disse deltakerne. Ingen gruppeforskjeller angående demografiske data ble funnet.
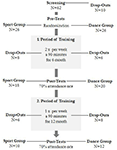
Figur 1. Flytskjema for deltakernes rekruttering.
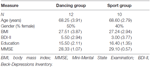
Tabell 1. Demografisk informasjon om deltakerne ved baseline.
Intervensjoner
intervensjonene ble delt inn i to perioder. I den første perioden trente fagene to ganger i uken i 90-min økter i 6 måneder. Av praktiske årsaker (tilgjengelighet av deltakere og trenere) besto den andre 12-måneders intervensjonsperioden av en redusert treningsfrekvens på en gang per uke. Begge intervensjonsprogrammene ble utført i en gruppesammenheng med musikk for å kontrollere psykososiale interaksjoner. Betinget belastning ble undersøkt ved å registrere pulsverdiene under treningsøktene og ved å beregne den enkelte treningspuls i Henhold Til Karvonen et al. (1957) med en faktor 0,6 for omfattende utholdenhetstrening. Derfor var de to treningsprogrammene sammenlignbare med hensyn til intensitet, varighet og frekvens. Begge ble overvåket av erfarne instruktører.
Dansegruppe
Deltakere i dansegruppen deltok på et nyutviklet treningsprogram der de hele tiden ble bedt om å lære nye bevegelsessekvenser. Disse koreografiene krevde koordinering av ulike kroppsdeler (dvs. ben, armer, trunk) i rom under forskjellige belastningsforhold (fysisk belastning, presisjon, situasjon og tidstrykk). Fagene måtte lære koreografiene av hjertet, og dermed stille høye krav til minne også. Programmet besto av fem forskjellige sjangere (line dance, jazz dance, rock » n » roll og square dance), som ble byttet etter hver fjerde økt. I løpet av intervensjonen ble koordinative krav og tidspress økt ved å innføre mer komplekse dansetrinn og koreografier og ved å øke slag per minutt i musikken.
Sport Group
Deltakerne i sportsgruppen fullførte et konvensjonelt treningsprogram for styrke og utholdenhet med hovedsakelig repeterende øvelser og lave krav til koordinering og hukommelse i hele kroppen. Hver økt besto av 20-min enheter av utholdenhet, styrke-utholdenhet og fleksibilitet trening. Utholdenhetstreningen ble utført på ergometre. I styrke-utholdenhetsenheten ble alternerende bevegelser (f. eks. biceps krøller, knebøy, sit-ups) utført, men komplekse helkroppsbevegelser ble unngått for å holde koordinative krav lave. Fleksibilitetsenheten besto hovedsakelig av strekkøvelser.
Utfallsmål
Kardiovaskulær Kondisjon
Kardiovaskulær kondisjon ble vurdert ved Fysisk Arbeidskapasitet 130 Test (PWC 130). PWC 130 er effekten (målt i watt) som et emne kan oppnå på et ergometer under en hjertefrekvens på 130 bpm. For beregninger brukte vi hvilepuls og relativ fysisk kapasitet .
Nevropsykologisk Testing
et omfattende batteri av nevropsykologiske tester ble utført på individene. For formålet med den nåværende studien rapporteres kun resultatene av en verbal kort – og langsiktig minnetest, «VLMT» (en tilpasset tysk versjon av «Rey Auditory Verbal Learning Test»; Helmstaedter Og Durwen, 1990) og et oppmerksomhetstestbatteri (Test Of Attentional Performance (TAP); Zimmermann og Fimm, 2002).
BDNF
Fastende blodprøver ble tatt om morgenen av de nevropsykologiske vurderingene. Fra blodprøvene ble plasmakonsentrasjonene av BDNF bestemt av sandwich ELISAs (BDNF DuoSets; r&D Systems, Wiesbaden, Tyskland) som tidligere beskrevet (Schega Et al., 2016).
MR
MR-bilder ble kjøpt på En 3 Tesla Siemens MAGNETOM Verio (Syngo MR B17) ved hjelp av en 32-kanals hodespole. Høyoppløselige t1-vektede MPRAGE-sekvenser ble anskaffet ved HJELP AV en 3d-magnetiseringsforberedt hurtiggradientekko-bildeprotokoll (224 sagittale skiver, voxelstørrelse: 0.8 × 0.8 × 0.8 mm3, TR: 2500 ms, TE: 3.47 ms, TI: 1100 ms, flippvinkel: 7°). MR-bildene ble analysert ved hjelp av voxel-basert morfometri (VBM) implementert I SPM 12 (Welcome Department Of Cognitive Neurology, London, UK). VBM er en hel-hjerne objektiv teknikk for analyse av regionale grå materie volum og vev endringer (Ashburner og Friston, 2000).Forbehandling involvert grå-materie segmentering, mal opprettelse via DARTEL, romlig normalisering til standardisert Montreal Neurological Institute (MNI) plass og utjevning Med En Gaussisk kjerne av 8 mm full bredde ved halv maksimum (FWHM).
for å analysere forskjellen i grå materie volumendringer mellom grupper, ble en fullfaktoriell design med faktorgruppen (dans, sport) og tid (0, 6 og 18 måneder) brukt. I tilfelle av signifikante gruppesamtaler , post hoc t-tester mellom påfølgende par av tidspunkter (0 vs. 6; 0 vs. 18; 6 vs. 18 måneder) ble beregnet separat for hver gruppe. En terskel for p < 0,001 (ukorrigert) ble brukt for alle analyser.
Resultater
presentasjonen av resultatene er strukturert som følger. Vi så først etter en generell intervensjonseffekt( faktortid), så etter differensielle effekter av de to tiltakene (interaksjonsgruppe × tid); til slutt ble en mer detaljert analyse av tidsdynamikken utført via post hoc parvise sammenligninger.
Kardiovaskulær Kondisjon
Kardiovaskulær kondisjon målt ved pwc 130 (Tabell 2, relativ fysisk kapasitet) var ikke forskjellig mellom gruppene ved baseline. VIDERE økte IKKE pwc 130-poengsummen betydelig i løpet av noen av intervensjonene.

Tabell 2. Midler og (SD) for fitness, kognitiv funksjon, bdnf plasmanivåer og totalt grått materievolum innen treningsgrupper over intervensjonen.
Nevropsykologiske Tester
en signifikant hovedeffekt av tid ble observert for verbal korttidshukommelse (VLMT tidlig tilbakekalling; F(2,19) = 6,438, p = 0,004, η2 = 0,253), verbal langsiktig fri tilbakekalling (VLMT sen tilbakekalling; F(2,19) = 3,387, p = 0,049, η2 = 0,244), verbal lang-term recognition (vlmt recognition; f(2,19) = 5.352, p = 0.009, ④2 = 0.220) OG ATTENTION REACTION TIME (subtest Fleksibilitet; f(2,19) = 19.156, p< 0.001, η2 = 0.489). Det oppsto ingen signifikante tidsmessige gruppeinteraksjoner.Post hoc parvise sammenligninger viste signifikante forbedringer fra baseline til 18 måneder og fra 6 til 18 måneder i ALLE TRE VLMT underkategorier i begge grupper. Når det gjelder TAP-reaksjonstider, ble det sett en signifikant forbedring i sammenligningen av baseline til 18-måneders data i begge grupper.
BDNF
Plasmanivåer AV BDNF ble analysert i blodprøver før treningsstart samt 6 måneder og 18 måneder etter treningsstart. Absolutte plasmanivåer AV bdnf er oppsummert i Tabell 2. De intraindividuelle endringene i BDNF-nivået viste en signifikant økning i dansegruppen, mens det intraindividuelle bdnf-nivået holdt seg konstant etter 6 måneders trening i sportsgruppen. Ingen ytterligere endringer i BDNF forekom etter 18 måneder i noen gruppe (Figur 2).
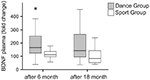
Figur 2. Intraindividuelle endringer I bdnf plasmanivåer etter intervensjon. Bdnf plasmanivåer ble analysert i blodprøver av deltakere som utførte et dansetreningsprogram eller et sportstreningsprogram før treningen begynte, etter 6 måneders trening og etter 18 måneders trening. Den relative økningen I BDNF-nivåer ble kvantifisert. Bdnf-nivåene økte signifikant i dansegruppen etter 6 måneders trening (Mann-Whitney U-test, p < 0,004) og falt nesten til baseline av forbehandlingsverdien etter 18 måneders dansetrening. Det var ingen endring over hele tidskurset i sportgruppen (Friedman test, p = 0,319), mens en signifikant endring over hele tidskurset ble observert i dansegruppen (Friedman test, p = 0,028). Boks tomter: minimum, 25. persentil, median, 75. persentil. * p ≤ 0.05.
MRI
en signifikant gruppe × tid interaksjon ble observert i venstre precentral gyrus og høyre parahippocampal. Post hoc t-tester mellom baseline og 6-måneders data viste en signifikant økning i grå materie volum i venstre precentral gyrus av bare dansere. I sammenligningen av baseline og 18-måneders data i tillegg til precentral gyrus, viste danserne en signifikant gråmassevolumøkning i høyre parahippocampal gyrus, som også var den eneste signifikante endringen i intervallet fra 6 til 18 måneder. Dermed oppstod volumøkningen i precentral gyrus etter 6 måneder og holdt seg stabil over det gjenværende dansetreningsintervallet, mens endringen i parahippocampal gyrus bare skjedde under det senere treningsintervallet (Figur 3).

Figur 3. Tid etter gruppeinteraksjonsanalyse, testing for større volumendringer i dansen sammenlignet med sportsgruppen. En signifikant økning i grå materie ble funnet i precentral gyrus (Montreal Neurological Institute(MNI) – koordinater: x = -16; y = -18; z = 77) og i parahippocampal gyrus gyrus (MNI-koordinater: x = 34; y = -26; z = -20). Boksen tomter viser de relative grå materie endringer i toppen voxel. * p ≤ 0.05.
Diskusjon
i denne studien sammenlignet vi effekten av deltakelse i enten et danseprogram eller et konvensjonelt fysisk treningsprogram på hjernefunksjon og volum hos friske eldre. Danseprogrammet var en nyutviklet intervensjon som krevde stadig å lære nye dansekoreografier. Det konvensjonelle sportsprogrammet fokuserte hovedsakelig på repeterende motorøvelser. Som et hovedfunn observerte vi at etter 6 måneders trening hadde volumene i dansernes venstre precentral gyrus økt mer enn de i sportsgruppen. Etter ytterligere 12 måneders trening ble det observert en ekstra volumøkning i dansernes høyre parahippocampal gyrus. BDNF-nivåene økte i løpet av de første 6 månedene av dansetrening og gikk tilbake til forbehandlingsverdiene etter 18 måneder. I den konvensjonelle sportsgruppen var en lignende økning I BDNF ikke tydelig. Fordi de kardiovaskulære treningsnivåene i løpet av intervensjonene holdt seg konstant i begge gruppene, kunne de observerte effektene ikke tilskrives forbedringer i fysisk kondisjon, men i stedet syntes å være relatert til de spesifikke egenskapene til danseprogrammet. Disse funksjonene inkluderte kravet om stadig å lære nye koreografier (dvs.minne), å integrere multisensorisk informasjon, å koordinere hele kroppen og å navigere i rommet.
Hjerneendringer
den precentrale gyrus er avgjørende for kontroll av frivillige motorfunksjoner. Økningen i grå materie volum i precentral gyrus i dansegruppen kan derfor ha vært basert på de komplekse og stadig skiftende bevegelsesmønstre som danserne måtte utføre. Disse bevegelsene krevde samtidig koordinering av flere deler av kroppen i forskjellige retninger og tilpasning til musikkens varierende rytmer (polycentrisk og polyrytmisk). Reflektere over disse komplekse koordinasjonskrav, Brown et al. (2006) har rapportert dans-induserte aktiveringer i putamen, den primære motor cortex og supplementary motor area (SMA), som VIST AV PET. Andre studier har indikert en sammenheng mellom koordineringskrav (f.eks. balansering, sjonglering) og neuroplasticitet i precentralområdet (Boyke et al., 2008; Taubert et al., 2010). Derfor var den danserelaterte volumøkningen i dette området i samsvar med forventninger basert på litteraturen.parahippocampal gyrus er en del av den ytre buen i det limbiske systemet og spiller en viktig rolle i arbeidsminne og episodisk minnehenting (Pantel et al., 2003). Ifølge Bliss og Lomo (1973) utgjør parahippocampal gyrus grensesnittet mellom minne og opplevelsesbevisstheten til nåtiden, fordi den er sammenkoblet av perforantkanalen både til regioner i frontal lobe, som er knyttet til arbeidsminne, og til hippocampus, den sentrale strukturen i episodisk minnekoding og romlig navigasjon. Mange vbm studier har rapportert en aldersrelatert volum tap i parahippocampal regioner (Tisserand et al., 2002). Videre Echur@varri et al. (2011) har antydet at parahippocampal atrofi er en tidlig biomarkør AV AD. Observasjonen som engasjerer seg i et danseprogram i en lengre periode, kan indusere neuroplastiske prosesser i denne viktige minnesregionen, derfor spesielt oppmuntrende når det gjelder å utvikle forebyggingsstrategier.
Temporal Dynamikk Av Grå Materie Hjernens Plastisitet
det observerte volumet øker i de to hjernegruppene utviklet på forskjellige tidspunkter. Dans førte til en volumøkning i motorområdene etter 6 måneder (Se Også Rehfeld et al., 2016), som holdt seg stabil i løpet av de påfølgende 12 månedene. Den danseassosierte volumøkningen i parahippocampal gyrus dukket opp senere og ble bare observert i 18-månedersdataene. De forskjellige temporale dynamikken i utviklingen av de to hjernegruppene kan være relatert til forskjeller i de underliggende cellulære mekanismene. Dyreforskning har antydet at angiogenese og dannelse av nye dendriter (Thomas Et al., 2012) forekommer raskt, mens endringer i neuropil forekommer mye langsommere (Black et al., 1990). Hos mennesker har raske økninger i gråstoffvolum i prefrontale regioner blitt observert etter bare 2 uker med motorlæring hos yngre voksne (Taubert et al., 2010). For å indusere neuroplasticitet i hippocampus, har lengre treningsperioder blitt anbefalt (Erickson et al., 2012; Niemann et al., 2014).
Interessant, de nevnte volumøkninger fra vår studie er i kontrast Til resultatene Av H ③nggi et al. (2010), som viste at profesjonelle ballettdansere har redusert grå materie volumer i venstre premotor cortex, SMA, Putamen og overlegen frontal gyrus og resultatene Av Hü et al. (2011), som rapporterer reduserte volumer i flere hjernegrupper, inkludert den fremre hippocampalformasjonen hos profesjonelle dansere og slackliner sammenlignet med ikke-profesjonelle. Derimot, disse studiene var tverrsnittsobservasjonsstudier som sammenlignet hjernen til profesjonelle dansere til de av ikke-profesjonelle. I tillegg, vår dansegruppe besto av gamle alderen nybegynnere. Det har vist seg at læring av en ny ferdighet først fører til rekruttering av ekstra nevrale ressurser. Senere, når ferdigheten blir mer automatisk, er det behov for mindre nevronressurser som kan føre til volumnedgang hos de med lang erfaring. Basert på disse innledende forholdene (nybegynnere, eldre voksne) og vårt spesialdesignede dansetreningsprogram, som krevde stadig ny læring av bevegelsesmønstre, er det mulig at effekter av spesialiseringsinducert volum reduseres som rapportert Av Hä Et al. (2010) er ikke nevnt.Det er generelt antatt at motor trening i utgangspunktet induserer hjernevolum øker. Langvarig trening fører imidlertid til automatisering, noe som kan ha motsatte effekter på kortikalt volum, fordi mindre kortikal kontroll er nødvendig etter at motorens ferdigheter er fullt etablert (Taube, 2008). Vårt dansetreningsprogram ble derfor spesielt designet for å unngå slik automatisering, noe som kan forklare hvorfor, i hvert fall innen 18 måneder, ble det ikke observert noen kortikale volumreduksjoner i vår studie.Kognitive funksjoner viste også ikke-lineær utvikling, hvorved verbalt minne bare økte i løpet av den andre treningsperioden. Når det gjelder oppmerksomhetsytelse, var betydelige forbedringer observerbare etter bare 6 måneder i begge grupper (Rehfeld et al., 2016). Disse funnene støtter tidligere rapporter om de gunstige effektene av fysiske inngrep på nevropsykologiske tester (Bamadis et al., 2014). Men i kognitive evnedata, i motsetning til hjernedataene, oppsto ingen gruppeforskjeller. Andre har rapportert overlegne effekter av kombinert kognitiv og fysisk trening i motsetning til enkeltintervensjoner (Oswald et al., 2006). Vi vil utvide våre tiltak ytterligere for å teste om gruppeforskjeller i kognisjon kan dukke opp på enda senere tidspunkter.
Underliggende Cellulære Og Molekylære Mekanismer Av Grå Materie Plastisitet
SELV OM VBM er en avbildningsmodalitet som avslører volumendringer i hjernen, tillater denne teknikken ikke kausale konklusjoner om de underliggende nevrofysiologiske prosessene. Neurogenese, synaptogenese og angiogenese er bare noen av mekanismene som har blitt foreslått å være grunnlaget for hjernevolumendringer (Zatorre et al., 2012). Som mediatorer av effekten av kardiovaskulær kondisjon på hjernen, studeres vekstfaktorer SOM BDNF, insulinlignende vekstfaktor (IGF) og nervevekstfaktor (NGF) (Kirk-Sanchez og McGough, 2014). Men i vår studie, i motsetning til tidligere (Erickson et al., 2011; Maass et al., 2015), var det ingen forskjeller i kondisjon mellom gruppene, og kondisjonsnivået endret seg ikke under intervensjonene. Sistnevnte observasjon var sannsynligvis relatert til vår kontroll over den enkelte hjertefrekvens, som vi hadde som mål å opprettholde i aerob sone. BDNF-endringer har imidlertid også vært forbundet med fysisk aktivitet, sosial interaksjon og positivt stress (Mattson, 2008), og ikke alle studier har observert EN ØKNING I BDNF etter kardiovaskulær trening (Vital et al., 2014). Endelig har dyreforskning antydet at koordinering, men ikke utholdenhetstrening, induserer synaptogenese og glialendringer (Black Et al., 1990). Sammen kan de nevnte tilleggsfaktorene som driver bdnf-sekresjon ha vært mer avgjørende under dans enn under treningsaktiviteter, og dermed forklare hvorfor bare dansere viste EN bdnf-økning i de første 6 månedene. Observasjonen om AT bdnf-nivåene gikk tilbake til baseline i de følgende 12 månedene, mens volumøkninger ble observert samtidig i parahippocampal gyrus, indikerer imidlertid at det må være andre faktorer involvert i voksen hjerneplastisitet enn de som representeres av bdnf-nivåer i perifert blod.
Angående nevrobiologiske mekanismer for utøvd-indusert plastisitet også begrepet hjerne reserve (Satz et al. 2011) bør vurderes. Konseptet med hjernereserve beskriver individuelle forskjeller i økt baseline adaptiv neuroplasticitet, noe som gir større dynamisk kapasitet for remodeling kortikale kretser til forskjellige stressorer (Barulli og Stern, 2013; Freret et al., 2015).
Perspektiver
resultatene av vår studie tyder på at en langsiktig dansintervensjon kan være bedre enn repeterende fysisk trening for å indusere neuroplasticitet i den aldrende menneskelige hjerne. Vi antar at denne fordelen er relatert til dansens multimodale natur, som kombinerer fysiske, kognitive og koordinative utfordringer. Så vidt vi vet er dette den første langsgående, randomiserte studien som anbefaler danseprogrammer som et middel for å forhindre grå materie og kognitiv nedgang hos eldre. Videre forskning er nødvendig for å avklare i større detalj den tidsmessige dynamikken og de underliggende nevrobiologiske mekanismene for dansinducert neuroplasticitet, og om denne intervensjonen virkelig har potensial til å redusere risikoen for nevrodegenerative sykdommer som Alzheimers.
Forfatterbidrag
PM designet OG utførte forskningen, analyserte dataene, skrev artikkelen. KR designet og utførte forskningen. MS analyserte dataene. AH og VL designet forskningen. MD: datainnsamling, artikkel revisjon. TB og JK designet forskningen, analyserte dataene. NGM designet forskningen, skrev artikkelen.
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
Ashburner, J. Og Friston, K. J. (2000). Voxel – basert morfometri-metodene. Neuroimage 11, 805-821. doi: 10.1006 / nimg.2000.0582
PubMed Abstract | CrossRef Full Text/Google Scholar
Bamadis, P. D., Vivas, A. B., Styliadis, C., Frantzidis, M., Klados, M., Schlee, W., et al. (2014). En gjennomgang av fysiske og kognitive tiltak i aldring. Neurosci. Biobehav. Åp 44, 206-220. doi: 10.1016 / j.neubiorev.2014.03.019
PubMed Abstract / CrossRef Full Text / Google Scholar
Barulli, D., Og Stern, Y. (2013). Effektivitet, kapasitet, kompensasjon, vedlikehold, plastisitet: nye konsepter i cognitice reserve. Trender Cogn. Sci. 17, 502–509. doi: 10.1016 / j. tics.2013.08.012
PubMed Abstract | CrossRef Full Text/Google Scholar
Black, J. E., Isaacs, K. R., Anderson, B. J., Alcantara, A. A., Og Greenough, W. T. (1990). Læring forårsaker synaptogenese, mens motoraktivitet forårsaker angiogenese, i cerebellar cortex av voksne rotter. Proc. Natl. Acad. Sci. U S A 87, 5568-5572. doi: 10.1073 / pnas.87.14.5568
PubMed Abstract / CrossRef Full Text / Google Scholar
Bliss, T. V., Og Lomo, T. (1973). Langvarig potensiering av synaptisk overføring i dentatet til den aneastetiserte kaninen etter stimulering av perforantbanen. J. Physiol. 232, 331–356. doi: 10.1113 / jfysiol.1973.sp010273
PubMed Abstract | CrossRef Full Text/Google Scholar
Boyke, J., Driemeyer, J., Gaser, C., Bü, C., Og Mai, A. (2008). Trening-indusert hjernestruktur endringer hos eldre. J. Neurosci. 28, 7031–7035. doi: 10.1523 / JNEUROSCI.0742-08.2008
PubMed Abstract | CrossRef Full Text/Google Scholar
Brun, S., Martinez, M., Og Parsons, M. (2006). Det neuronale grunnlaget for menneskelig dans. Cereb. Cortex 16, 1157-1167. doi: 10.1093/cercor/bhj057
PubMed Abstract | CrossRef Full Text | Google Scholar
Colcombe, S. J., Erickson, K. I., Scalf, P. E., Kim, J. S., Prakash, R., McAuley, E., Et al. (2006). Aerob trening øker hjernevolumet i aldrende mennesker. J. Gerontol. A. Biol. Sci. Med. Sci. 61, 1166–1170. doi: 10.1093 / gerona / 61.11.1166
PubMed Abstract | CrossRef Full Text/Google Scholar
Echupunvarri, C., Aalten, P., Uyling, H. B. M., Jacobs, H. I. L., Visser, P. J., Gronenschild, M., et al. (2011). Atrofi i parahippocampal gyrus som en tidlig biomarkør Av Alzheimers sykdom. Brain Struct. Funct. 215, 265–271. doi: 10.1007 / s00429-010-0283-8
PubMed Abstract / CrossRef Full Text/Google Scholar
Edelmann, E., Lessmann, V., Og Brigadski, T. (2014). Pre – og postsynaptiske vendinger i bdnf-sekresjon og virkning i synaptisk plastisitet. Nevrofarmakologi 76, 610-627. doi: 10.1016 / j.neuropharm.2013.05.043
PubMed Abstract | CrossRef Full Text/Google Scholar
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., et al. (2011). Trening øker størrelsen på hippocampus og forbedrer hukommelsen. Proc. Natl. Acad. Sci. USA 108, 3017-3022. doi: 10.1073 / pnas.1015950108
PubMed Abstract / CrossRef Full Text / Google Scholar
Erickson, K. I., Weinstein, A. M., Og Lopez, O. L. (2012). Fysisk aktivitet, hjerneplastisitet og Alzheimers sykdom. Arch. Med. Res. 43, 615-621. doi: 10.1016 / j.arcmed.2012.09.008
PubMed Abstract | CrossRef Full Text / Google Scholar
Flö, A., Ruscheweyh, R., Krü, K., Willemer, C., Vinter, B., Vö, K., et al. (2010). Fysisk aktivitet og minnefunksjoner: er neurotrofiner og cerebral grå materie volum den manglende lenken? Neuroimage 49, 2756-2763. doi: 10.1016 / j. neuroimage.2009.10.043
PubMed Abstract | CrossRef Full Text/Google Scholar
Freret, T., Gaudreau, P., Schumann-Bard, P., Billard, Jm, Og Popa-Wagner, a. (2015). Mekanismer som ligger til grunn for den nevrobeskyttende effekten av hjernereserve mot sen depresjon. J. Neural. Transm. (Wien) 122, S55-S61. doi: 10.1007 / s00702-013-1154-2
PubMed Abstract | CrossRef Full Text/Google Scholar
Gregory, S. M., Parker, B., Og Thompson, P. D. (2012). Fysisk aktivitet, kognitiv funksjon og hjernehelse: hva er treningsopplæringens rolle i forebygging av demens? Brain Sci. 2, 684–708. doi: 10.3390 / brainsci2040684
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
Hä, Bezzola, L., og Jä, L. (2010). Strukturell neuroplasticitet i sensorimotorisk nettverk av profesjonelle kvinnelige ballettdansere. Hum. Brain Mapp. 31, 1196–1206. doi: 10.1002 / hbm.20928
PubMed Abstract | CrossRef Full Text/Google Scholar
Helmstaedter, C., Og Durwen, H. F. (1990). Verbal læring og retensjon test. Et nyttig og differensiert verktøy for å evaluere verbal minneytelse. Schweiz. Arch. Neurol. Psychiatr. (1985) 141, 21–30.
PubMed Abstract/Google Scholar
Huang, E. J., Og Reichardt, L. F. (2001). Neurotrofiner: roller i nevronutvikling og funksjon. Annu. Pastor Neurosci. 24, 677–736. doi: 10.1146 / annurev.nevro.24.1.677
PubMed Abstract / CrossRef Full Text/Google Scholar
Hü, K., Binetti, C., Hamilton, D. A., Stephan, T., Flanagin, V. L., Linn, J., et al. (2011). Strukturell og funksjonell plastisitet av hippocampalformasjonen i profesjonelle dansere og slackliner. Hippocampus 21, 855-865. doi: 10.1002 / hipo.20801
PubMed Abstract / CrossRef Full Text / Google Scholar
Karvonen, M. J., Kentala, E., Og Mustala, O. (1957). Effekten av trening på hjertefrekvens: en longitudinell studie. Anne. Med. Exp. Biol. Fenn. 35, 307–315.
PubMed Abstract/Google Scholar
Kattenstroth, J. C., Kalisch, T., Holt, S., Tegethoff, M., Og Dinse, H. R. (2013). Seks måneders danseintervensjon forbedrer postural, sensorimotorisk og kognitiv ytelse hos eldre uten å påvirke kardio-respiratoriske funksjoner. Front. Aldring Neurosci. 5:5. doi: 10.3389 / fnagi.2013.00005
PubMed Abstract / CrossRef Full Text/Google Scholar
Kattenstroth, J. C., Kolankowska, I., Kalisch, T., Og Dinse, H. R. (2010). Overlegen sensorisk, motorisk og kognitiv ytelse hos eldre personer med flerårige dansaktiviteter. Front. Aldring Neurosci. 2:31. doi: 10.3389 / fnagi.2010.00031
PubMed Abstract / CrossRef Full Text / Google Scholar
Kempermann, G., Kuhn, H. G., Og Gage, F. H. (1997). Flere hippocampale nevroner hos voksne mus som lever i et beriket miljø. Natur. 386, 493–495. doi: 10.1038/386493a0
PubMed Abstract / CrossRef Full Text / Google Scholar
Kirk-Sanchez, Nj, Og McGough, E. L. (2014). Fysisk trening og kognitiv ytelse hos eldre: nåværende perspektiver. Clin. Interv. Aldring 9, 51-62. doi: 10.2147 / CIA.S39506
PubMed Abstract / CrossRef Full Text / Google Scholar
Lessmann, V., Og Brigadski, T. (2009). Mekanismer, steder og kinetikk av synaptisk bdnf-sekresjon: en oppdatering. Neurosci. Res. 65, 11-22. doi: 10.1016 / j.neures.2009.06.004
PubMed Abstract | CrossRef Full Text/Google Scholar
Maass, A., Dü, S., Goerke, M., Becke, A., Sobieray, U., Neumann, K., Et al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M., Hö, a., Kaufmann, J., et al. (2016). Effekter av dans og helse sport trening på den grå hjernen saken av friske eldre. Sportwiss. 46, 213-222. doi: 10.1007 / s12662-016-0411-6
CrossRef Full Text / Google Scholar
Niemann, C., Godde, B., Og Voelcker-Rehage, C. (2014). Ikke bare kardiovaskulær, men også koordinert trening øker hippocampalvolumet hos eldre voksne. Front. Aldring Neurosci. 6:170. doi: 10.3389 / fnagi.2014.00170
PubMed Abstract / CrossRef Full Text/Google Scholar
Oswald, W. D., Gunzelmann, T., Rupprecht, R., Og Hagen, B. (2006). Differensielle effekter av enkelt versus kombinert kognitiv og fysisk trening med eldre voksne: SimA-studien i et 5-årig perspektiv. Eur. J. Aldring 3, 179-192. doi: 10.1007 / s10433-006-0035-Z
CrossRef Full Text | Google Scholar
Pantel, J., Kratz, B., Essig, M., Og Schrö, J. (2003). Parahippocampal volum underskudd hos personer med aldring-assosiert kognitiv nedgang. Er. J. Psykiatri 160, 379-382. doi: 10.1176 / appi.ajp.160.2.379
PubMed Abstrakt | CrossRef Fulltekst/Google Scholar
Park, H., Og Poo, M. M. (2013). Neurotrofin regulering av nevrale kretsutvikling og funksjon. Nat. Pastor Neurosci. 14, 7–23. doi: 10.1038/nrn3379
PubMed Abstract | CrossRef Full Text | Google Scholar
Rehfeld, K., H@kelmann, A., Kaufmann, J., Og Mü, N. G. (2016). «Effekter av en dans vs. en treningsopplæring på hjernens plastisitet, balanseytelse og oppmerksomhet hos sunne seniorer: en ny tilnærming MED SPM 12 for parvis longitudinell gruppe sammenligning,» In Proceeding-International Conference Of Sports And Neuroscience (Magdeburg), 127-138.I 2011 ble Det opprettet En egen organisasjon. Hjerne og kognitiv reserve: mediator (er) og konstruktiv validitet, en kritikk. J. Clin. Exp. Neuropsykol. 33, 121–130. doi: 10.1080/13803395.2010.493151
PubMed Abstract | CrossRef Full Text | Google Scholar
Schega, L., Peter, B., Brigadski, T., Leß, V., Isermann, B., Hamacher, D., et al. (2016). Effekt av intermitterende normobarisk hypoksi på aerob kapasitet og kognitiv funksjon hos eldre mennesker. J. Sci. Med. Sport 19, 941-945. doi: 10.1016/j. jsams.2016.02.012
PubMed Abstract / CrossRef Full Text/Google Scholar
Taube, W. (2008). Nevrofysiologiske tilpasninger som svar på balansetrening. Dtsch. Z. Sportmed. 63, 273–277. doi: 10.5960 / dzsm.2012.030
CrossRef Full Text/Google Scholar
Taubert, M., Draganski, B., Anwander, A., Mü, K., Horstmann, A., Villringer, A., Et al. (2010). Dynamiske egenskaper av menneskelig hjernestruktur: læringsrelaterte endringer i kortikale områder og tilhørende fiberforbindelser. J. Neurosci. 30, 11670–11677. doi: 10.1523 / JNEUROSCI.2567-10.2010
PubMed Abstract | CrossRef Full Text/Google Scholar
Thomas, A. G., Dennis, A., Bandettini, P. a., Og Johansen-Berg, H. (2012). Effekten av aerob aktivitet på hjernestrukturen. Front. Psychol. 3:86. doi: 10.3389 / fpsyg.2012.00086
PubMed Abstract | CrossRef Full Text/Google Scholar
Tisserand, D. J., Pruessner, J. C., Sanz Arigita, E. J., van Boxtel, M. P., Evans, A. C., Jolles, J., et al. (2002). Regionale frontale kortikale volumer reduseres differensielt i aldring: EN MR-studie for å sammenligne volumetriske tilnærminger og voxelbasert morfometri. Neuroimage 17, 657-669. doi: 10.1016 / s1053-8119 (02)91173-0
PubMed Abstract | CrossRef Full Text/Google Scholar
van Praag, H., Shubert, T., Zhao, C., og Gage, F. H. (2005). Trening forbedrer læring og hippocampal neurogenese hos eldre mus. J. Neurosci. 25, 8680–8685. doi: 10.1523 / JNEUROSCI.1731-05. 2005
PubMed Abstract / CrossRef Full Text/Google Scholar
Verghese, J., Lipton, R. B., Katz, M. J., Hall, C. B., Dervy, C. A., Kuslansky, G., et al. (2003). Fritidsaktiviteter og risiko for demens hos eldre. N. Engl. J. Med. 348, 2508–2516. doi: 10.1056/NEJMoa022252
PubMed Abstract | CrossRef Full Text | Google Scholar
Vital, T. M., Stein, A. M., De Melo Coelho, F. G., Arantes, F. J., Teodorov, E., Santos-Galduró, R. F., Et al. (2014). Fysisk trening og vaskulær endotelial vekstfaktor (VEGF) hos eldre: en systematisk oversikt. Arch. Gerontol. Geriatr. 59, 234–239. doi: 10.1016 / j.archger.2014.04.011
PubMed Abstract | CrossRef Full Text/Google Scholar
Voelcker-Rehage, C. Og Niemann, C. (2013). Strukturelle og funksjonelle hjerneendringer relatert til ulike typer fysisk aktivitet over hele levetiden. Neurosci. Biobehav. Åp 37, 2268-2295. doi: 10.1016 / j.neubiorev.2013.01.028
PubMed Abstract | CrossRef Full Text/Google Scholar
Zatorre, R. J., Fields, R. D., Og Johansen-Berg, H. (2012). Plastisitet i grått og hvitt: neuroimaging endringer i hjernestruktur under læring. Nat. Neurosci. 15, 528–536. doi: 10.1038 / nn.3045
PubMed Abstract / CrossRef Full Text/Google Scholar
Zimmermann, P., Og Fimm, B. (2002). Testbatteri for oppmerksomhetstesting (TAP), versjon 1.7, manuell-Part1. Wü: Psytest.