
Introducción
El cambio demográfico actual en las sociedades occidentales, que implica un aumento relativo y absoluto en el número de personas mayores, ha despertado un creciente interés científico en temas geriátricos. En este contexto, los conceptos de envejecimiento exitoso son cada vez más importantes. Varios estudios han indicado que el ejercicio físico puede desempeñar un papel clave en el envejecimiento saludable y en la prevención del deterioro cognitivo y las enfermedades neurodegenerativas (Colcombe et al., 2006; Erickson et al., 2011). Revisiones recientes que resumen estudios epidemiológicos, transversales e intervencionistas apoyan la actividad física como un método propicio para inducir neuroplasticidad a finales de la edad adulta (Gregory et al., 2012; Voelcker-Rehage y Niemann, 2013; Bamadis et al., 2014). Erickson et al. (2012) han llegado a la conclusión de que la aptitud cardiorrespiratoria y los niveles de actividad física aeróbica más altos están asociados con volúmenes de materia gris más grandes en las regiones prefrontales y el hipocampo. Sin embargo, no solo la aptitud cardiovascular, sino también el ejercicio coordinativo (Niemann et al., 2014) y entrenamiento cognitivo (Bamadis et al., 2014) han demostrado inducir la plasticidad de la materia gris y mejorar las funciones cognitivas en adultos mayores.
La investigación en animales ha sugerido que una combinación de ejercicio físico con enriquecimiento sensorial tiene el efecto más fuerte en la génesis de nuevas neuronas, predominantemente en el hipocampo, y que solo esta combinación garantiza la supervivencia duradera de las células recién nacidas (Kempermann et al., 1997; van Praag et al., 2005). Kattenstroth et al. (2013) han sugerido que «las actividades de baile deben considerarse como un equivalente de las condiciones ambientales enriquecidas para los seres humanos, ya que proporcionan a un individuo mayores demandas sensoriales, motoras y cognitivas.»A pesar de esta afirmación alentadora, los estudios que examinan los efectos del entrenamiento de danza en la estructura o función cerebral son escasos. Después de una intervención de baile de 6 meses, Kattenstroth et al. (2010) han reportado mejoras significativas en el rendimiento cognitivo, táctil y motor en las personas mayores participantes. Los resultados de un estudio prospectivo de 469 sujetos mayores de 75 años durante un período de seguimiento medio de 5,1 años han indicado que la danza se asocia con un riesgo notablemente reducido de demencia (Verghese et al., 2003). Sin embargo, Hüfner et al. (2011) han reportado volúmenes reducidos en varias regiones cerebrales, incluida la formación del hipocampo anterior y partes de la corteza vestibular parieto-insular en bailarines profesionales y slackliners en comparación con no profesionales. Este hallazgo puede indicar que el entrenamiento intensivo y repetitivo de las mismas habilidades motoras conduce en el contexto de la especialización a volúmenes reducidos de algunas regiones cerebrales. En este estudio, por lo tanto, diseñamos un programa especial de entrenamiento de danza que requería constantemente que los participantes aprendieran nuevos patrones de movimiento (Müller et al., 2016). Para evaluar los beneficios específicos de esta intervención, comparamos nuestro programa de baile de nuevo diseño con un grupo de control activo en lugar de pasivo, que participó en un programa de acondicionamiento físico deportivo de salud convencional en el que los participantes generalmente realizaban ejercicios físicos repetitivos, como andar en bicicleta en un ergómetro. Además, como nos interesaba la dinámica temporal de las intervenciones, evaluamos los efectos en la estructura y función del cerebro después de 6 y 18 meses de entrenamiento. Al hacerlo, buscamos evaluar si es beneficioso extender las intervenciones, porque algunas regiones cerebrales pueden requerir más entrenamiento que otras, o si hay un límite después del cual más entrenamiento se vuelve perjudicial. Finalmente, en busca de un mecanismo potencial subyacente a la neuroplasticidad, medimos los niveles de BDNF en la sangre periférica. Varios estudios han sugerido que el BDNF promueve la diferenciación de nuevas neuronas y sinapsis (Huang y Reichardt, 2001; Lessmann y Brigadski, 2009; Park y Poo, 2013; Edelmann et al., 2014). Por lo tanto, se ha propuesto que el BDNF sea un mediador de la neuroplasticidad en adultos (Flöel et al., 2010).
Materiales y métodos
Participantes y Diseño Experimental
El estudio fue diseñado como una intervención controlada de 18 meses. El estudio fue aprobado por el comité de ética de la Universidad Otto-von-Guericke, Magdeburgo, y todos los sujetos firmaron un escrito informado y no recibieron pago por su participación. Sesenta y dos ancianos sanos (63-80 años) reclutados a través de anuncios en los periódicos locales fueron seleccionados para el estudio. Los criterios de exclusión fueron claustrofobia, tinnitus, implantes metálicos, tatuajes, diabetes mellitus, depresión (Inventario de Depresiones de Beck, BDI-II > 13), déficits cognitivos (Miniexamen del Estado Mental, MMSE < 27), enfermedades neurológicas y ejercicio regular (≥1 h/semana). Sobre la base de estos criterios, se excluyeron 10 sujetos. Los 52 participantes restantes se asignaron aleatoriamente al grupo de baile o deporte utilizando el sitio web www.randomization.com y controlando la edad, el estado MMSE y la condición física. Las evaluaciones se realizaron al inicio, después de 6 y 18 meses de entrenamiento (Figura 1). Veintidós participantes completaron toda la intervención y todas las mediciones. En el cuadro 1 se proporcionan datos demográficos detallados de estos participantes. No se encontraron diferencias de grupo con respecto a los datos demográficos.
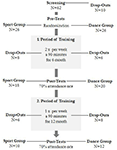
la Figura 1. Diagrama de flujo de la contratación de participantes.
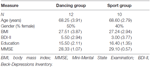
la Tabla 1. Información demográfica de los participantes al inicio del estudio.
Intervenciones
Las intervenciones fueron separados en dos períodos. En el primer período, los sujetos entrenaron dos veces por semana en sesiones de 90 minutos durante 6 meses. Por razones prácticas (disponibilidad de participantes e instructores), el segundo período de intervención de 12 meses comprendió una frecuencia de capacitación reducida de una vez por semana. Ambos programas de intervención se realizaron en un contexto grupal con música para controlar las interacciones psicosociales. La carga condicional se examinó registrando los valores de pulso durante las sesiones de entrenamiento y calculando la frecuencia cardíaca de entrenamiento individual según Karvonen et al. (1957) con un factor de 0,6 para un entrenamiento de resistencia extensivo. Por lo tanto, los dos programas de entrenamiento fueron comparables en términos de intensidad, duración y frecuencia. Ambos fueron supervisados por instructores experimentados.
Grupo de Baile
Los participantes en el grupo de baile asistieron a un programa de entrenamiento de nuevo diseño en el que se les pedía constantemente que aprendieran nuevas secuencias de movimiento. Estas coreografías requerían la coordinación de diferentes partes del cuerpo (es decir, piernas, brazos, tronco) en el espacio bajo diferentes condiciones de tensión (tensión física, precisión, situación y presión de tiempo). Los sujetos tenían que aprender las coreografías de memoria, imponiendo así altas exigencias a la memoria también. El programa comprendía cinco géneros diferentes (line dance, jazz dance, rock «n» roll y square dance), que se cambiaban después de cada cuarta sesión. A lo largo de la intervención, las demandas de coordinación y la presión de tiempo se incrementaron mediante la introducción de movimientos de baile y coreografías más complejas y el aumento de los latidos por minuto en la música.
Grupo Deportivo
Los participantes del grupo deportivo completaron un programa de entrenamiento de fuerza y resistencia convencional con ejercicios principalmente repetitivos y bajas demandas en términos de coordinación y memoria de todo el cuerpo. Cada sesión comprendía unidades de 20 minutos de entrenamiento de resistencia, resistencia y flexibilidad. El entrenamiento de resistencia se realizó en cicloergómetros. En la unidad de fuerza-resistencia, se realizaron movimientos alternos (por ejemplo, rizos de bíceps, sentadillas, abdominales), pero se evitaron movimientos complejos de todo el cuerpo para mantener bajas las demandas de coordinación. La unidad de flexibilidad consistía principalmente en ejercicios de estiramiento.
Medidas de resultados
Aptitud Cardiovascular
La aptitud cardiovascular se evaluó mediante la Prueba de Capacidad Física de Trabajo 130 (PWC 130). PWC 130 es la potencia de salida (medida en vatios) que un sujeto es capaz de lograr en un cicloergómetro con una frecuencia cardíaca de 130 lpm. Para los cálculos, se utilizó la frecuencia cardíaca en reposo y la capacidad física relativa .
Pruebas neuropsicológicas
Se realizó una extensa batería de pruebas neuropsicológicas a los sujetos. Para el propósito del presente estudio, solo se reportan los resultados de una prueba de memoria verbal a corto y largo plazo, la «VLMT» (una versión adaptada en alemán de la «Prueba de Aprendizaje Verbal Auditivo Rey»; Helmstaedter y Durwen, 1990) y una batería de prueba de atención (Prueba de Rendimiento Atencional (TAP); Zimmermann y Fimm, 2002).
BDNF
Se tomaron muestras de sangre en ayunas en las mañanas de las evaluaciones neuropsicológicas. A partir de las muestras de sangre, las concentraciones plasmáticas de BDNF se determinaron mediante ELISAs sándwich (DuoSets de BDNF; R&D Systems, Wiesbaden, Alemania) como se describió anteriormente (Schega et al., 2016).
Las imágenes de RMN
se obtuvieron en un Siemens MAGNETOM Verio de 3 Tesla (Syngo MR B17) utilizando una bobina de cabeza de 32 canales. Las secuencias de MPRAGE ponderadas en T1 de alta resolución se adquirieron utilizando un protocolo de imágenes de eco de gradiente rápido preparado para magnetización 3D (224 cortes sagitales, tamaño de voxel: 0,8 × 0,8 × 0,8 mm3, TR: 2500 ms, TE: 3,47 ms, TI: 1100 ms, ángulo de giro: 7°). Las imágenes de RM se analizaron mediante morfometría basada en vóxeles (MBV) implementada en SPM 12 (Welcome Department of Cognitive Neurology, Londres, Reino Unido). La MBV es una técnica imparcial de todo el cerebro para el análisis del volumen regional de materia gris y los cambios en los tejidos (Ashburner y Friston, 2000).
El preprocesamiento incluyó segmentación de materia gris, creación de plantillas a través de DARTEL, normalización espacial al espacio estandarizado del Instituto Neurológico de Montreal (MNI) y suavizado con un núcleo gaussiano de 8 mm de ancho completo a la mitad del máximo (FWHM).
Para analizar la diferencia en los cambios de volumen de materia gris entre grupos, se aplicó un diseño factorial completo con el grupo de factores (danza, deporte) y el tiempo (0, 6 y 18 meses). En el caso de interacciones significativas de grupo × tiempo, pruebas t post hoc entre pares consecutivos de puntos de tiempo (0 vs. 6; 0 vs. 18; 6 vs. 18 meses) se calcularon por separado para cada grupo. Se aplicó un umbral de p < 0,001 (no corregido) para todos los análisis.
Resultados
La presentación de los resultados se estructura de la siguiente manera. Primero buscamos un efecto de intervención general (factor tiempo), luego buscamos los efectos diferenciales de las dos intervenciones (grupo de interacción × tiempo); finalmente, se realizó un análisis más detallado de la dinámica temporal a través de comparaciones de pares post hoc.
Aptitud cardiovascular
La aptitud cardiovascular medida por el PWC 130 (Tabla 2, capacidad física relativa) no difirió entre los grupos al inicio del estudio. Además, la puntuación de PWC 130 no aumentó significativamente durante el transcurso del tiempo de ninguna de las intervenciones.

la Tabla 2. Medios y (SD) para la aptitud, el funcionamiento cognitivo, los niveles plasmáticos de BDNF y el volumen total de materia gris dentro de los grupos de entrenamiento durante la intervención.
Pruebas neuropsicológicas
Se observó un efecto principal significativo del tiempo para la memoria verbal a corto plazo (memoria temprana VLMT; F(2,19) = 6,438, p = 0,004, η2 = 0,253), memoria libre verbal a largo plazo (memoria tardía VLMT; F(2,19) = 3,387, p = 0,049, η2 = 0,244), memoria verbal a largo plazo-reconocimiento de términos (reconocimiento VLMT; F(2,19) = 5,352, p = 0,009, η2 = 0,220) y tiempo de reacción de atención (flexibilidad de pruebas secundarias; F(2,19) = 19,156, p< 0,001, η2 = 0,489). No se observaron interacciones de grupo significativas en el tiempo.
Las comparaciones post hoc por parejas mostraron mejoras significativas desde el inicio hasta los 18 meses y de los 6 a los 18 meses en las tres subcategorías de TMLV en ambos grupos. En cuanto a los tiempos de reacción del TAP, se observó una mejora significativa en la comparación de los datos basales con los de 18 meses en ambos grupos.
BDNF
Los niveles plasmáticos de BDNF se analizaron en muestras de sangre antes del inicio del entrenamiento, así como a los 6 y 18 meses después del inicio del entrenamiento. Los niveles plasmáticos absolutos de BDNF se resumen en la Tabla 2. Los cambios intraindividuales en el nivel de BDNF revelaron un aumento significativo en el grupo de baile, mientras que el nivel de BDNF intraindividual se mantuvo constante después de 6 meses de entrenamiento en el grupo deportivo. No se produjeron más cambios en el BDNF después de 18 meses en ningún grupo (Figura 2).
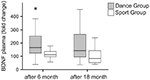
la Figura 2. Cambios intraindividuales en los niveles plasmáticos de BDNF después de la intervención. Los niveles plasmáticos de BDNF se analizaron en muestras de sangre de participantes que realizaban un programa de entrenamiento de baile o un programa de entrenamiento deportivo antes del inicio del entrenamiento, después de 6 meses de entrenamiento y después de 18 meses de entrenamiento. Se cuantificó el aumento relativo de los niveles de BDNF. Los niveles de BDNF aumentaron significativamente en el grupo de baile después de 6 meses de entrenamiento (prueba U de Mann-Whitney, p < 0,004) y disminuyeron casi hasta el valor basal del pretratamiento después de 18 meses de entrenamiento de baile. En el grupo deportivo (prueba de Friedman, p = 0,319) no hubo cambio durante todo el curso del tiempo, mientras que en el grupo de baile se observó un cambio significativo durante todo el curso del tiempo (prueba de Friedman, p = 0,028). Parcelas de caja: mínimo, percentil 25, mediana, percentil 75. * p ≤ 0,05.
RM
Se observó una interacción significativa de grupo × tiempo en el giro precentral izquierdo y el parahippocampal derecho. Las pruebas t post hoc entre los datos basales y los de 6 meses mostraron un aumento significativo en el volumen de materia gris en el giro precentral izquierdo de solo las bailarinas. En la comparación de los datos basales y de 18 meses, además del giro precentral, los bailarines mostraron un aumento significativo del volumen de materia gris en el giro parahipocampal derecho, que también fue el único cambio significativo en el intervalo de 6 a 18 meses. Por lo tanto, el aumento de volumen en el giro precentral surgió después de 6 meses y se mantuvo estable durante el intervalo de entrenamiento de baile restante, mientras que el cambio en el giro parahippocampal ocurrió solo durante el intervalo de entrenamiento posterior (Figura 3).
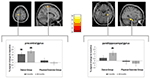
Figura 3. Análisis de interacción de tiempo por grupo, probando cambios de volumen mayores en el baile en comparación con el grupo deportivo. Se encontró un aumento significativo de la materia gris en el giro precentral (coordenadas del Instituto Neurológico de Montreal (MNI): x = -16; y = -18; z = 77) y en el giro parahippocampal (coordenadas MNI: x = 34; y = -26; z = -20). Las gráficas de caja muestran los cambios relativos de la materia gris en el voxel de pico. * p ≤ 0,05.
Discusión
En este estudio, comparamos los efectos de la participación en un programa de baile o en un programa deportivo de fitness físico convencional sobre la función y el volumen del cerebro en personas mayores sanas. El programa de baile fue una intervención de nuevo diseño que requería aprender constantemente nuevas coreografías de baile. El programa deportivo convencional se centró principalmente en ejercicios motores repetitivos. Como hallazgo principal, observamos que después de 6 meses de entrenamiento, los volúmenes en el giro precentral izquierdo de los bailarines habían aumentado más que los del grupo deportivo. Después de otros 12 meses de entrenamiento, se observó un aumento de volumen adicional en el giro parahippocampal derecho de los bailarines. Los niveles de BDNF aumentaron durante los primeros 6 meses de entrenamiento de baile y volvieron a los valores previos al tratamiento después de 18 meses. En el grupo de deportes convencionales, no se observó un aumento similar del BDNF. Debido a que los niveles de aptitud cardiovascular durante el transcurso de las intervenciones se mantuvieron constantes en ambos grupos, los efectos observados no pudieron atribuirse a mejoras en la aptitud física, sino que parecían estar relacionados con las características específicas del programa de danza. Estas características incluían el requisito de aprender constantemente nuevas coreografías (es decir, memoria), integrar información multisensorial, coordinar todo el cuerpo y navegar en el espacio.
Cambios cerebrales
El giro precentral es esencial para el control de las funciones motoras voluntarias. El aumento del volumen de materia gris en el giro precentral en el grupo de baile puede, por lo tanto, haberse basado en los patrones de movimiento complejos y en constante cambio que los bailarines tenían que realizar. Estos movimientos requerían la coordinación simultánea de varias partes del cuerpo en diferentes direcciones y el ajuste a los ritmos variables de la música (policéntricos y polirrítmicos). Reflexionando sobre estos complejos requisitos de coordinación, Brown et al. (2006) han reportado activaciones inducidas por la danza en el putamen, la corteza motora primaria y el área motora suplementaria (SMA), como se muestra en la PET. Otros estudios han indicado una asociación entre las demandas de coordinación (por ejemplo, equilibrio, malabarismo) y la neuroplasticidad en la región precentral (Boyke et al., 2008; Taubert et al., 2010). Por lo tanto, el aumento de volumen relacionado con la danza en esta área fue consistente con las expectativas basadas en la literatura.
El giro parahippocampal es parte del arco exterior del sistema límbico y juega un papel importante en la memoria de trabajo y la recuperación de memoria episódica(Pantel et al., 2003). Según Bliss y Lomo (1973), el giro parahippocampal constituye la interfaz entre la memoria y la conciencia experiencial del presente, porque está interconectado por el tracto perforante tanto a las regiones del lóbulo frontal, que están asociadas con la memoria de trabajo, como al hipocampo, la estructura central en la codificación de la memoria episódica y la navegación espacial. Muchos estudios de MBV han reportado una pérdida de volumen relacionada con la edad en regiones parahipocampales (Tisserand et al., 2002). Además, Echávarri et al. (2011) han sugerido que la atrofia parahippocampal es un biomarcador temprano de la EA. La observación de que participar en un programa de danza durante un período más largo puede inducir procesos neuroplásicos en esta región crucial de la memoria, por lo tanto, es particularmente alentadora en términos de desarrollar estrategias de prevención.
Dinámica Temporal de la Plasticidad Cerebral de la Materia Gris
El volumen observado aumenta en las dos regiones cerebrales desarrolladas en momentos diferentes. El baile llevó a un aumento de volumen en las áreas motoras después de 6 meses (ver también Rehfeld et al., 2016), que se mantuvo estable durante los siguientes 12 meses. El aumento de volumen asociado a la danza en el giro parahippocampal surgió más tarde y se observó solo en los datos de 18 meses. Las diferentes dinámicas temporales en la evolución de las dos regiones cerebrales pueden estar relacionadas con diferencias en los mecanismos celulares subyacentes. La investigación en animales ha sugerido que la angiogénesis y la formación de nuevas dendritas (Thomas et al., 2012) ocurren rápidamente, mientras que los cambios en el neuropilo ocurren mucho más lentamente (Black et al., 1990). En humanos, se han observado aumentos rápidos en el volumen de materia gris en las regiones prefrontales después de solo 2 semanas de aprendizaje motor en adultos más jóvenes (Taubert et al., 2010). Para inducir neuroplasticidad en el hipocampo, se han recomendado períodos de entrenamiento más largos (Erickson et al., 2012; Niemann et al., 2014).
Curiosamente, los aumentos de volumen mencionados en nuestro estudio contrastan con los resultados de Hänggi et al. (2010), que reveló que los bailarines de ballet profesionales han disminuido los volúmenes de materia gris en la corteza premotora izquierda, AME, Putamen y circunvolución frontal superior y los resultados de Hüfner et al. (2011), que reportan volúmenes reducidos en varias regiones cerebrales, incluida la formación del hipocampo anterior en bailarines profesionales y slackliners en comparación con los no profesionales. Sin embargo, estos estudios fueron estudios observacionales transversales que compararon los cerebros de bailarines profesionales con los de no profesionales. Además, nuestro grupo de baile estaba formado por novatos ancianos. Se ha demostrado que el aprendizaje de una nueva habilidad conduce primero al reclutamiento de recursos neuronales adicionales. Más tarde, cuando la habilidad se vuelve más automática, se necesitan menos recursos neuronales, lo que puede conducir a disminuciones de volumen en aquellos con experiencia a largo plazo. En base a estas condiciones iniciales (novatos, adultos mayores) y nuestro programa de entrenamiento de baile diseñado especialmente, que requería un aprendizaje constantemente nuevo de los patrones de movimiento, es posible que los efectos de la especialización inducida por el volumen disminuyan, según lo informado por Hänggi et al. (2010) no se observan.
Generalmente se piensa que el entrenamiento motor inicialmente induce aumentos de volumen cerebral. Sin embargo, el entrenamiento prolongado conduce a la automatización, que puede tener los efectos opuestos en el volumen cortical, porque se necesita menos control cortical después de que las habilidades motoras se hayan establecido completamente (Taube, 2008). Nuestro programa de entrenamiento de baile, por lo tanto, fue especialmente diseñado para evitar dicha automatización, lo que puede explicar por qué, al menos en 18 meses, no se observaron disminuciones de volumen cortical en nuestro estudio.
Las funciones cognitivas también mostraron un desarrollo no lineal, por lo que la memoria verbal aumentó solo durante el segundo período de entrenamiento. En cuanto al rendimiento de la atención, se observaron mejoras significativas después de solo 6 meses en ambos grupos (Rehfeld et al., 2016). Estos hallazgos apoyan informes previos sobre los efectos beneficiosos de las intervenciones físicas en pruebas neuropsicológicas (Bamadis et al., 2014). Sin embargo, en los datos de la capacidad cognitiva, en contraste con los datos del cerebro, no surgieron diferencias de grupo. Otros han reportado efectos superiores del entrenamiento cognitivo y físico combinado en comparación con intervenciones individuales (Oswald et al., 2006). Ampliaremos aún más nuestras intervenciones para probar si las diferencias grupales en la cognición podrían surgir incluso en momentos posteriores.
Mecanismos Celulares y Moleculares Subyacentes de la Plasticidad de la Materia Gris
Aunque la MBV es una modalidad de imagen que revela cambios de volumen en el cerebro, esta técnica no permite conclusiones causales con respecto a los procesos neurofisiológicos subyacentes. La neurogénesis, la sinaptogénesis y la angiogénesis son solo algunos de los mecanismos que se han sugerido que son la base de los cambios de volumen cerebral (Zatorre et al., 2012). Como mediadores de los efectos de la aptitud cardiovascular en el cerebro, se están estudiando factores de crecimiento como el BDNF, el factor de crecimiento similar a la insulina (IGF) y el factor de crecimiento nervioso (NGF) (Kirk-Sanchez y McGough, 2014). Sin embargo, en nuestro estudio, en contraste con los anteriores (Erickson et al., 2011; Maass et al., 2015), no hubo diferencias en la aptitud cardiovascular entre los grupos y los niveles de aptitud física no cambiaron durante las intervenciones. Esta última observación probablemente se relacionó con nuestro control de la frecuencia cardíaca individual, que pretendíamos mantener en la zona aeróbica. Sin embargo, los cambios en el BDNF también se han asociado con la actividad física, la interacción social y el estrés positivo (Mattson, 2008), y no todos los estudios han observado un aumento del BDNF después del entrenamiento cardiovascular (Vital et al., 2014). Finalmente, la investigación en animales ha sugerido que la coordinación, pero no el entrenamiento de resistencia, induce sinaptogénesis y cambios gliales (Black et al., 1990). Juntos, los factores adicionales nombrados que impulsan la secreción de BDNF pueden haber sido más cruciales durante el baile que durante las actividades de acondicionamiento físico, lo que explica por qué solo los bailarines mostraron un aumento de BDNF en los primeros 6 meses. Sin embargo, la observación de que los niveles de BDNF volvieron a los valores basales en los 12 meses siguientes, mientras que los aumentos de volumen se observaron simultáneamente en la circunvolución parahipocampal, indica que debe haber otros factores involucrados en la plasticidad cerebral adulta que los representados por los niveles de BDNF en la sangre periférica.
En cuanto a los mecanismos neurobiológicos de la plasticidad inducida por el ejercicio, también el concepto de reserva cerebral (Satz et al., 2011). El concepto de reserva cerebral describe diferencias individuales en un aumento de la neuroplasticidad adaptativa basal, que proporciona una mayor capacidad dinámica para remodelar los circuitos corticales a diferentes factores de estrés (Barulli y Stern, 2013; Freret et al., 2015).
Perspectivas
Los resultados de nuestro estudio sugieren que una intervención de baile a largo plazo podría ser superior al ejercicio físico repetitivo para inducir la neuroplasticidad en el cerebro humano envejecido. Suponemos que esta ventaja está relacionada con la naturaleza multimodal de la danza, que combina desafíos físicos, cognitivos y coordinativos. Hasta donde sabemos, este es el primer estudio longitudinal aleatorizado que recomienda programas de baile como medio para prevenir la materia gris y el deterioro cognitivo en los ancianos. Se necesita más investigación para aclarar con mayor detalle la dinámica temporal y los mecanismos neurobiológicos subyacentes de la neuroplasticidad inducida por la danza y si esta intervención realmente tiene el potencial de reducir el riesgo de enfermedades neurodegenerativas como el Alzheimer.
Contribuciones de los autores
PM diseñó y realizó la investigación, analizó los datos, escribió el artículo. KR diseñó y realizó la investigación. MS analizó los datos. AH y VL diseñaron la investigación. MD: recopilación de datos, revisión de artículos. TB y JK diseñaron la investigación, analizaron los datos. NGM diseñó la investigación, escribió el artículo.
Declaración de Conflicto de Intereses
Los autores declaran que la investigación se realizó en ausencia de relaciones comerciales o financieras que pudieran interpretarse como un posible conflicto de intereses.
Ashburner, J., and Friston, K. J. (2000). Morfometría basada en vóxeles – los métodos. Neuroimagen 11, 805-821. doi: 10.1006 / nimg.2000.0582
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Bamadis, P. D., Vivas, A. B., Styliadis, C., Frantzidis, M., Klados, M., Schlee, W., et al. (2014). Una revisión de las intervenciones físicas y cognitivas en el envejecimiento. Neurociencia. Biobehav. Apo. 44, 206-220. doi: 10.1016 / j. neubiorev.2014.03.019
Resumen de PubMed / Texto completo Cruzado / Google Scholar
Barulli, D., and Stern, Y. (2013). Eficiencia, capacidad, compensación, mantenimiento, plasticidad: conceptos emergentes en reserva cognitiva. Trends Cogn. Sci. 17, 502–509. doi: 10.1016 / j. tics.2013.08.012
Resumen de PubMed / Texto completo Cruzado/Google Scholar
Black, J. E., Isaacs, K. R., Anderson, B. J., Alcantara, A. A., and Greenough, W. T. (1990). El aprendizaje causa sinaptogénesis, mientras que la actividad motora causa angiogénesis, en la corteza cerebelosa de ratas adultas. Proc. Natl. Acad. Sci. U S A 87, 5568-5572. doi: 10.1073 / pnas.87.14.5568
Resumen de PubMed / Texto completo de CrossRef/Google Scholar
Bliss, T. V., and Lomo, T. (1973). Potenciación de larga duración de la transmisión sináptica en el dentado del conejo aneoestetizado tras la estimulación de la vía perforante. J. Physiol. 232, 331–356. doi: 10.1113 / jfisiol.1973.sp010273
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Boyke, J., Driemeyer, J., Gaser, C., Büchel, C., y Puede, A. (2008). Cambios en la estructura cerebral inducidos por el entrenamiento en los ancianos. J. Neurocirujano. 28, 7031–7035. doi: 10.1523 / JNEUROSCI.0742-08.2008
Resumen de PubMed | Texto completo cruzado/Google Scholar
Brown, S., Martinez, M., and Parsons, M. (2006). La base neuronal de la danza humana. Cereb. Cortex 16, 1157-1167. doi: 10.1093/cercor/bhj057
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Colcombe, S. J., Erickson, K. I., Scalf, P. E., Kim, J. S., Prakash, R., McAuley, E., et al. (2006). El entrenamiento aeróbico aumenta el volumen cerebral en los seres humanos de edad avanzada. J. Gerontol. A. Biol. Sci. Mediterráneo. Sci. 61, 1166–1170. doi: 10.1093 / gerona / 61.11.1166
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Echávarri, C., Aalten, P., Uyling, H. B. M., Jacobs, H. I. L., Visser, P. J., Gronenschild, M., et al. (2011). Atrofia en el giro parahipocampal como biomarcador temprano de la enfermedad de Alzheimer. Estructura Cerebral. Funt. 215, 265–271. doi: 10.1007 / s00429-010-0283-8
Resumen de PubMed / Texto completo cruzado / Google Scholar
Edelmann, E., Lessmann, V. y Brigadski, T. (2014). Giros pre y postsinápticos en la secreción de BDNF y acción en la plasticidad sináptica. Neurofarmacología 76, 610-627. doi: 10.1016 / j. neuropharm.2013.05.043
Resumen de PubMed / Texto completo Cruzado/Google Scholar
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., et al. (2011). El entrenamiento físico aumenta el tamaño del hipocampo y mejora la memoria. Proc. Natl. Acad. Sci. U S A 108, 3017-3022. doi: 10.1073 / pnas.1015950108
Resumen de PubMed / Texto completo cruzado / Google Scholar
Erickson, K. I., Weinstein, A. M., and Lopez, O. L. (2012). Actividad física, plasticidad cerebral y enfermedad de Alzheimer. Arco. Mediterráneo. Res. 43, 615-621. doi: 10.1016 / j. arcmed.2012.09.008
Resumen de PubMed | Texto completo Cruzado/Google Scholar
Flöel, A., Ruscheweyh, R., Krüger, K., Willemer, C., Winter, B., Völker, K., et al. (2010). Actividad física y funciones de memoria: ¿las neurotrofinas y el volumen de materia gris cerebral son el eslabón perdido? Neuroimage 49, 2756-2763. doi: 10.1016 / j.neuroimagen.2009.10.043
Resumen de PubMed / Texto completo Cruzado/Google Scholar
Freret, T., Gaudreau, P., Schumann-Bard, P., Billard, J. M., and Popa-Wagner, A. (2015). Mecanismos subyacentes al efecto neuroprotector de la reserva cerebral contra la depresión tardía de la vida. J. Neural. Transm. (Viena) 122, S55 a S61. doi: 10.1007 / s00702-013-1154-2
Resumen de PubMed / Texto completo cruzado / Google Scholar
Gregory, S. M., Parker, B., and Thompson, P. D. (2012). Actividad física, función cognitiva y salud cerebral: ¿cuál es el papel del entrenamiento físico en la prevención de la demencia? Sci Cerebral. 2, 684–708. doi: 10.3390/brainsci2040684
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Hänggi, J., Koeneke, S., Bezzola, L., and Jäncke, L. (2010). Neuroplasticidad estructural en la red sensomotora de bailarinas de ballet profesionales. Zumbido. Mapa Cerebral. 31, 1196–1206. doi: 10.1002 / hbm.20928
Resumen de PubMed | Texto completo cruzado | Google Scholar
Helmstaedter, C., y Durwen, H. F. (1990). La prueba de aprendizaje y retención verbal. Una herramienta útil y diferenciada para evaluar el rendimiento de la memoria verbal. Schweiz. Arco. Neurol. Psiquiatras. (1985) 141, 21–30.
PubMed Abstract/Google Scholar
Huang, E. J., y Reichardt, L. F. (2001). Neurotrofinas: roles en el desarrollo y la función neuronal. Annu. Reverendo Neurocirujano. 24, 677–736. doi: 10.1146 / annurev.neuro.24.1.677
Resumen de PubMed / Texto completo Cruzado/Google Scholar
Hüfner, K., Binetti, C., Hamilton, D. A., Stephan, T., Flanagin, V. L., Linn, J., et al. (2011). Plasticidad estructural y funcional de la formación del hipocampo en bailarines profesionales y slackliners. Hipocampo 21, 855-865. doi: 10.1002 / hipo.20801
Resumen de PubMed / Texto completo Cruzado / Google Scholar
Karvonen, M. J., Kentala, E., and Mustala, O. (1957). The effects of training on heart rate: a longitudinal study (en inglés). Ana. Mediterráneo. Exp. Biol. Fenn. 35, 307–315.
Resumen de PubMed/Google Scholar
Kattenstroth, J.C., Kalisch, T., Holt, S., Tegethoff, M., and Dinse, H. R. (2013). Seis meses de intervención de danza mejoran el rendimiento postural, sensomotor y cognitivo en ancianos sin afectar las funciones cardiorrespiratorias. Delantero. Neurociencia envejecida. 5:5. doi: 10.3389 / fnagi.2013.00005
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Kattenstroth, J. C., Kolankowska, I., Kalisch, T., and Dinse, H. R. (2010). Rendimiento sensorial, motor y cognitivo superior en personas mayores con actividades de baile de varios años. Delantero. Neurociencia envejecida. 2:31. doi: 10.3389 / fnagi.2010.00031
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Kempermann, G., Kuhn, H. G., y Gage, F. H. (1997). Más neuronas del hipocampo en ratones adultos que viven en un ambiente enriquecido. Naturaleza. 386, 493–495. doi: 10.1038 / 386493a0
Resumen de PubMed | Texto completo cruzado/Google Scholar
Kirk-Sanchez, N. J., and McGough, E. L. (2014). Ejercicio físico y rendimiento cognitivo en ancianos: perspectivas actuales. Clin. Interv. Envejeciendo 9, 51-62. doi: 10.2147 / CIA.S39506
Resumen de PubMed / Texto completo de CrossRef / Google Scholar
Lessmann, V., and Brigadski, T. (2009). Mechanisms, locations, and kinetics of synaptic BDNF secretion: an update (en inglés). Neurociencia. Res. 65, 11 a 22. doi: 10.1016 / j. neures.2009.06.004
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Maass, A., Düzel, S., Goerke, M., Becke, A., Sobieray, U., Neumann, K., et al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M., Hökelmann, A., Kaufmann, J., et al. (2016). Efectos del entrenamiento deportivo de danza y salud en la materia gris del cerebro de las personas mayores sanas. Sportwiss. 46, 213-222. doi: 10.1007 / s12662-016-0411-6
Texto completo cruzado / Google Scholar
Niemann, C., Godde, B., y Voelcker-Rehage, C. (2014). No solo el ejercicio cardiovascular, sino también el ejercicio coordinado aumenta el volumen del hipocampo en adultos mayores. Delantero. Neurociencia envejecida. 6:170. doi: 10.3389 / fnagi.2014.00170
PubMed Abstract | CrossRef Texto Completo / Google Scholar
Oswald, W. D., Gunzelmann, T., Rupprecht, R., y Hagen, B. (2006). Efectos diferenciales del entrenamiento cognitivo y físico individual versus combinado con adultos mayores: el estudio SimA en una perspectiva de 5 años. EUR. J. Ageing 3, 179-192. doi: 10.1007 / s10433-006-0035-z
CrossRef Full Text / Google Scholar
Pantel, J., Kratz, B., Essig, M., and Schröde, J. (2003). Déficits de volumen parahippocampal en sujetos con deterioro cognitivo asociado al envejecimiento. Ser. J. Psiquiatría 160, 379-382. doi: 10.1176 / appi.ajp.160.2.379
Resumen de PubMed / Texto completo de CrossRef/Google Scholar
Park, H., and Poo, M. M. (2013). Regulación de la neurotrofina del desarrollo y la función de los circuitos neuronales. NAT. Reverendo Neurocirujano. 14, 7–23. doi: 10.1038 / nrn3379
Resumen de PubMed / Texto completo cruzado/Google Scholar
Rehfeld, K., Hökelmann, A., Kaufmann, J., and Müller, N. G. (2016). «Effects of a dance vs.a fitness training on brain plasticity, balance performance and attention in healthy seniores: a new approach with SPM 12 for pairwise longitudinal group comparison», en Proceeding-International Conference of Sports and Neuroscience (Magdeburg), 127-138.
Satz, P., Cole, M. A., Hardy, D. J., and Rassovsky, Y. (2011). Cerebro y reserva cognitiva: mediador(es) y validez de constructo, una crítica. J. Clin. Exp. Neuropsicol. 33, 121–130. doi: 10.1080 | 13803395.2010.493151
Resumen de PubMed/Texto completo cruzado/Google Scholar
Schega, L., Peter, B., Brigadski, T., Leßmann, V., Isermann, B., Hamacher, D., et al. (2016). Efecto de la hipoxia normobárica intermitente sobre la capacidad aeróbica y la función cognitiva en personas mayores. J. Sci. Mediterráneo. Sport 19, 941-945. doi: 10.1016 / j. jsams.2016.02.012
Resumen de PubMed | Texto completo Cruzado/Google Scholar
Taube, W. (2008). Adaptaciones neurofisiológicas en respuesta al entrenamiento de equilibrio. Dtsch. Z. Sportmed. 63, 273–277. doi: 10.5960 / dzsm.2012.030
Texto completo de CrossRef | Google Scholar
Taubert, M., Draganski, B., Anwander, A., Müller, K., Horstmann, A., Villringer, A., et al. (2010). Propiedades dinámicas de la estructura del cerebro humano: aprendizaje de cambios relacionados en áreas corticales y conexiones de fibra asociadas. J. Neurocirujano. 30, 11670–11677. doi: 10.1523 / JNEUROSCI.2567-10.2010
Resumen de PubMed / Texto completo cruzado / Google Scholar
Thomas, A. G., Dennis, A., Bandettini, P. A., y Johansen-Berg, H. (2012). Los efectos de la actividad aeróbica en la estructura cerebral. Delantero. Psicol. 3:86. doi: 10.3389 / fpsyg.2012.00086
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Tisserand, D. J., Pruessner, J. C., Sanz Arigita, E. J., van Boxtel, M. P., Evans, A. C., Jolles, J., et al. (2002). Los volúmenes corticales frontales regionales disminuyen diferencialmente en el envejecimiento: un estudio de RM para comparar abordajes volumétricos y morfometría basada en vóxeles. Neuroimagen 17, 657-669. doi: 10.1016 / s1053-8119 (02)91173-0
Resumen PubMed | Texto completo cruzado | Google Scholar
van Praag, H., Shubert, T., Zhao, C., and Gage, F. H. (2005). El ejercicio mejora el aprendizaje y la neurogénesis del hipocampo en ratones de edad avanzada. J. Neurocirujano. 25, 8680–8685. doi: 10.1523 / JNEUROSCI.1731-05.2005
PubMed Abstract | CrossRef Texto Completo | Google Scholar
Verghese, J., Lipton, R. B., Katz, J. M., Hall, C. B., Dervy, C. A., Kuslansky, G., et al. (2003). Actividades de ocio y el riesgo de demencia en las personas mayores. N. Engl. J. Med. 348, 2508–2516. doi: 10.1056 / NEJMoa022252
Resumen de PubMed | Texto completo cruzado | Google Académico
Vital, T. M., Stein, A. M., de Melo Coelho, F. G., Arantes, F. J., Teodorov, E., Santos-Galduróz, R. F., et al. (2014). Ejercicio físico y factor de crecimiento endotelial vascular (VEGF) en ancianos: una revisión sistemática. Arco. Gerontol. Geriátrico. 59, 234–239. doi: 10.1016 / j. archger.2014.04.011
Resumen de PubMed / Texto completo Cruzado / Google Scholar
Voelcker-Rehage, C. y Niemann, C. (2013). Cambios cerebrales estructurales y funcionales relacionados con diferentes tipos de actividad física a lo largo de la vida. Neurociencia. Biobehav. Apo. 37, 2268-2295. doi: 10.1016 / j. neubiorev.2013.01.028
Resumen de PubMed / Texto completo cruzado / Google Scholar
Zatorre, R. J., Fields, R. D., and Johansen-Berg, H. (2012). Plasticidad en gris y blanco: cambios de neuroimagen en la estructura cerebral durante el aprendizaje. NAT. Neurociencia. 15, 528–536. doi: 10.1038 / nn.3045
Resumen de PubMed / Texto completo cruzado / Google Scholar
Zimmermann, P., and Fimm, B. (2002). Batería de prueba para pruebas de atención (TAP), versión 1.7, manual, Parte 1. Würselen: Psytest.