
Introdução
O atual mudança demográfica nas sociedades Ocidentais, com o envolvimento de um familiar e um aumento absoluto no número de pessoas mais velhas, tem suscitado crescente interesse científico em geriatria problemas. Neste contexto, os conceitos de envelhecimento bem sucedido estão se tornando cada vez mais importantes. Vários estudos indicaram que o exercício físico pode desempenhar um papel fundamental no envelhecimento saudável e na prevenção do declínio cognitivo e das doenças neurodegenerativas (Colcombe et al., 2006; Erickson et al., 2011). Recent reviews sumarizing epidemiological, cross-sectional and interventional studies support physical activity as a propitious method to induce neuroplasticity in late adulthood (Gregory et al., 2012; Voelcker-Rehage and Niemann, 2013; Bamadis et al., 2014). Erickson et al. (2012) concluíram que níveis mais elevados de aptidão cardiorrespiratória e de actividade física aeróbica estão associados a maiores volumes de matéria cinzenta nas regiões pré-frontal e hipocampo. No entanto, não só a aptidão cardiovascular, mas também o exercício coordenado (Niemann et al., 2014) e formação cognitiva (Bamadis et al., 2014) têm sido mostrados para induzir plasticidade da matéria cinzenta e melhorar as funções cognitivas em adultos mais velhos.a pesquisa Animal sugeriu que uma combinação de exercício físico com enriquecimento sensorial tem o efeito mais forte na gênese de novos neurônios—predominantemente no hipocampo—e que apenas esta combinação garante a sobrevivência duradoura das células recém-nascidas (Kempermann et al., 1997; van Praag et al., 2005). Kattenstroth et al. (2013) têm sugerido que “as atividades de dança devem ser consideradas como um equivalente a condições ambientais enriquecidas para os seres humanos, uma vez que eles fornecem a um indivíduo com maior exigência sensorial, motora e cognitiva.”Apesar desta afirmação encorajadora, estudos examinando os efeitos do treinamento de dança na estrutura ou função cerebral são escassos. Após uma intervenção de 6 meses de dança, Kattenstroth et al. (2010) relataram melhorias significativas no desempenho cognitivo, táctil e motor nos idosos participantes. Os resultados de um estudo prospectivo com 469 indivíduos com mais de 75 anos, num período mediano de acompanhamento de 5, 1 anos, indicaram que a dança está associada a um risco de demência marcadamente reduzido (Verghese et al., 2003). No entanto, Hüfner et al. (2011) têm relatado volumes reduzidos em várias regiões do cérebro, incluindo a formação hipocampal anterior e partes do córtex vestibular parieto-insular em dançarinos profissionais e slackliners em comparação com não profissionais. Esta constatação pode indicar que a formação intensiva e repetitiva das mesmas competências motoras conduz, no contexto da especialização, a volumes reduzidos de algumas regiões do cérebro. Neste estudo, projetamos, portanto, um programa especial de treinamento de dança que constantemente exigia que os participantes aprendessem novos padrões de movimento (Müller et al., 2016). Para avaliar os benefícios específicos desta intervenção, comparamos o nosso Programa de dança recentemente concebido a um grupo de controlo activo e não passivo, que participou num programa de fitness Desportivo de saúde convencional no qual os participantes tipicamente realizavam exercícios físicos repetitivos, tais como ciclismo num ergômetro. Além disso, como estávamos interessados na dinâmica temporal das intervenções, avaliamos os efeitos sobre a estrutura e funcionamento do cérebro após 6 e 18 meses de treinamento. Ao fazê-lo, procurámos avaliar se é benéfico alargar as intervenções, porque algumas regiões do cérebro podem necessitar de mais formação do que outras, ou se existe um limite após o qual mais formação se torna prejudicial. Finalmente, em busca de um mecanismo potencial subjacente à neuroplasticidade, medimos os níveis de BDNF no sangue periférico. Vários estudos têm sugerido que o BDNF promove a diferenciação de novos neurônios e sinapses (Huang e Reichardt, 2001; Lessmann e Brigadski, 2009; Park e Poo, 2013; Edelmann et al., 2014). BDNF, portanto, foi proposto para ser um mediador da neuroplasticidade adulta (Flöel et al., 2010).o estudo foi concebido como uma intervenção controlada de 18 meses. O estudo foi aprovado pelo Comitê de Ética da Universidade Otto-von-Guericke, Magdeburg, e todos os sujeitos assinaram um comunicado escrito e não receberam pagamento por sua participação. Sessenta e dois idosos saudáveis (63-80 anos) recrutados através de anúncios em jornais locais foram selecionados para o estudo. Os critérios de exclusão foram claustrofobia, zumbido, implantes metálicos, tatuagens, diabetes mellitus, depressão (Beck-Depressões Inventory, BDI-II > 13), déficit cognitivo (Mini-Exame do Estado Mental, MEEM < 27), doenças neurológicas e regular exercício (≥1 h/semana). Com base nestes critérios, foram excluídos 10 indivíduos. Os restantes 52 participantes foram então distribuídos aleatoriamente para o grupo de dança ou esporte usando o site www.randomization.com e controlando para a idade, o estado MMSE e a aptidão física. As avaliações foram realizadas no início, após 6 e 18 meses de formação (Figura 1). Vinte e dois participantes completaram toda a intervenção e todas as medições. O quadro 1 apresenta dados demográficos pormenorizados para estes participantes. Não foram encontradas diferenças de grupo em relação aos dados demográficos.
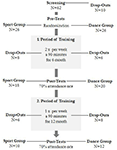
Figura 1. Fluxograma do recrutamento dos participantes.
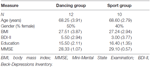
a Tabela 1. Informação demográfica sobre os participantes na linha de base.as intervenções foram separadas em dois períodos. No primeiro período, os indivíduos treinaram duas vezes por semana em sessões de 90 minutos durante 6 meses. Por razões práticas (disponibilidade de participantes e formadores), o segundo período de intervenção de 12 meses incluiu uma frequência de formação reduzida de uma vez por semana. Ambos os programas de intervenção foram realizados em um contexto de grupo com música para controlar as interações psicossociais. A carga condicional foi examinada gravando os valores dos impulsos durante as sessões de treino e calculando a frequência cardíaca individual de treino de acordo com Karvonen et al. (1957) with a factor 0.6 for extensive endurance training. Assim, os dois programas de treinamento eram comparáveis em termos de intensidade, duração e frequência. Ambos foram supervisionados por instrutores experientes.
grupo de dança
participantes no grupo de dança participaram de um programa de treinamento recém-projetado em que eles foram constantemente convidados a aprender novas sequências de movimento. Estas coreografias exigiam a coordenação de diferentes partes do corpo (isto é, pernas, braços, tronco) no espaço sob diferentes condições de tensão (tensão física, precisão, situação e pressão de tempo). Os sujeitos tiveram que aprender as coreografias de cor, impondo assim elevadas exigências à memória também. O programa compreendia cinco gêneros diferentes (dança em linha, jazz, rock “N” roll e square dance), que foram trocados após cada quarta sessão. Ao longo da intervenção, as exigências coordenadas e a pressão do tempo foram aumentadas pela introdução de movimentos de dança e coreografias mais complexos e pelo aumento das batidas por minuto na música.
Grupo Desportivo
participantes no Grupo Desportivo completaram um programa convencional de treino de resistência com exercícios repetitivos e baixas exigências em termos de coordenação de corpo inteiro e memória. Cada sessão compreendia 20 minutos de unidades de resistência, resistência e treino de flexibilidade. O treinamento de resistência foi realizado em ergómetros de ciclo. Na unidade força-resistência, movimentos alternados (por exemplo, curvas bíceps, agachamentos, abdominais) foram realizados, mas movimentos complexos de todo o corpo foram evitados para manter as demandas coordenadas baixas. A unidade de flexibilidade consistiu principalmente em exercícios de alongamento.a aptidão Cardiovascular foi avaliada pelo teste de capacidade física de trabalho 130 (PWC 130). PWC 130 é a potência de saída (medida em watts) que um sujeito é capaz de alcançar em um ergômetro de ciclo sob uma frequência cardíaca de 130 bpm. Para cálculos, usamos o ritmo cardíaco em repouso e a capacidade física relativa .testes neuropsicológicos foram realizados uma extensa bateria de testes neuropsicológicos nos indivíduos. Para o propósito do presente estudo, apenas os resultados de um verbal de curto e de longo prazo teste de memória, o “VLMT” (adaptação de uma versão alemã da “Rey Auditory Verbal Learning Test”; Helmstaedter e Durwen, 1990) e uma atenção bateria de testes (Teste de Desempenho Atencional (TAP); Zimmermann e Fimm, 2002) são relatados.
BDNF
amostras de sangue em jejum foram colhidas de manhã nas avaliações neuropsicológicas. A partir das amostras de sangue, as concentrações plasmáticas de BDNF foram determinadas por Elisas em sanduíche (DuoSets BDNF; R&d Systems, Wiesbaden, Alemanha), como descrito anteriormente (Schega et al., 2016).imagens MR foram adquiridas em um Tesla Siemens MAGNETOM Verio (Syngo MR B17) usando uma bobina de cabeça de 32 canais. Sequências de MPRAGE ponderadas em alta resolução T1 foram adquiridas usando um protocolo de imagem de eco de gradiente rápido preparado por magnetização 3D (224 fatias sagittais, tamanho voxel: 0,8 × 0,8 × 0,8 mm3, TR: 2500 ms, TE: 3.47 ms, TI: 1100 ms, ângulo flip: 7°). As imagens do MR foram analisadas usando morfometria baseada em voxel (VBM) implementada no SPM 12 (Welcome Department of Cognitive Neurology, Londres, Reino Unido). VBM é uma técnica de análise do volume de massa cinzenta regional e das alterações tecidulares (Ashburner e Friston, 2000).
pré-processamento envolveu segmentação de matéria cinzenta, criação de modelos através de DARTEL, normalização espacial para o espaço padronizado do Montreal Neurological Institute (MNI) e alisamento com um núcleo Gaussiano de 8 mm de largura completa a metade do máximo (FWHM).
para analisar a diferença de volume de matéria cinzenta entre grupos, foi aplicado um design totalmente factorial com o grupo de fatores (dança, esporte) e tempo (0, 6 e 18 meses). No caso de grupos significativos × interacções temporais, testes t post hoc entre pares consecutivos de pontos temporais (0 vs. 6; 0 vs. 18; 6 vs. 18 meses) foram calculados separadamente para cada grupo. Foi aplicado um limiar de p< 0, 001 (não corrigido) para todas as análises.
resultados
a apresentação dos resultados é estruturada da seguinte forma. Primeiramente buscamos um efeito de Intervenção Geral (tempo fator), depois buscamos efeitos diferenciais das duas intervenções (grupo de interação × tempo); finalmente, uma análise mais detalhada da dinâmica temporal foi realizada através de comparações post hoc emparelhadas.a aptidão Cardiovascular
a aptidão Cardiovascular medida pela PWC 130 (Tabela 2, capacidade física relativa) não diferiu entre os grupos basais. Além disso, a pontuação PwC 130 não aumentou significativamente ao longo do tempo de qualquer das intervenções.

Tabela 2. Meios e (DP) para a aptidão física, o funcionamento cognitivo, os níveis plasmáticos de BDNF e o volume total de matéria cinzenta nos grupos de treino durante a intervenção.
Testes Neuropsicológicos
Um efeito principal significativo do tempo foi observada para o verbal, memória de curto prazo (VLMT início de recall; F(2,19) = 6.438, p = 0,004, η2 = 0.253), verbal de longo prazo livre de recall (VLMT tarde recordar; F(2,19) = 3.387, p = 0.049, η2 = 0.244), verbal de longo prazo (reconhecimento VLMT de reconhecimento; F(2,19) = 5.352, p = 0,009, η2 = 0.220) atenção e tempo de reação (subtest flexibilidade; F(2,19) = 19.156, p < de 0,001, η2 = 0.489). Não surgiram interacções significativas entre o tempo e o grupo.as comparações Post hoc em pares mostraram melhorias significativas desde a linha de base até aos 18 meses e de 6 a 18 meses nas três subcategorias VLMT de ambos os grupos. No que respeita aos tempos de reacção da TAP, observou-se uma melhoria significativa na comparação da linha de base com os dados de 18 meses em ambos os grupos.os níveis plasmáticos de BDNF foram analisados em amostras de sangue antes do início do treino, bem como 6 meses e 18 meses após o início do treino. Os níveis plasmáticos absolutos de BDNF estão resumidos na Tabela 2. As alterações intra-individuais no nível de BDNF revelaram um aumento significativo no grupo de dança, enquanto o nível intra-individual de BDNF permaneceu constante após 6 meses de formação no Grupo Desportivo. Não ocorreram mais alterações no BDNF após 18 meses em nenhum grupo (Figura 2).
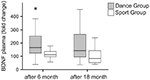
Figura 2. Alterações intra-individuais nos níveis plasmáticos de BDNF após intervenção. Os níveis plasmáticos de BDNF foram analisados em amostras de sangue de participantes executando um programa de treinamento de dança ou um programa de treinamento esportivo antes do início do treinamento, após 6 meses de treinamento e após 18 meses de treinamento. O aumento relativo dos níveis de BDNF foi quantificado. Os níveis de BDNF aumentaram significativamente no grupo de dança após 6 meses de treinamento (teste de Mann-Whitney U, p < 0.004) e declinou quase até o valor inicial do pré-tratamento após 18 meses de treinamento de dança. Não houve mudança ao longo de todo o tempo no sport group (teste Friedman, p = 0.319), enquanto uma mudança significativa ao longo de todo o tempo foi observada no grupo de dança (teste Friedman, p = 0.028). Box plots: mínimo, percentil 25, mediana, percentil 75. * p ≤ 0, 05.
MRI
foi observada interacção significativa entre o grupo × Tempo no giro pré-Central esquerdo e o parahippocampal direito. Os testes T Post hoc entre os dados de base e os de 6 meses mostraram um aumento significativo no volume de matéria cinzenta no giro pré-Central esquerdo de apenas os dançarinos. Na comparação da linha de base e os 18 meses de dados, além de o giro precentral, os bailarinos apresentaram um significativo aumento do volume de matéria cinzenta no direito parahippocampal giro, que também foi a única mudança significativa no intervalo de 6 a 18 meses. Assim, o aumento de volume no giro pré-Central surgiu após 6 meses e permaneceu estável durante o restante intervalo de treinamento de dança, enquanto a mudança no giro parahippocampal ocorreu apenas durante o intervalo de treinamento posterior (Figura 3).
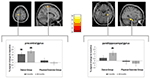
Figura 3. Tempo por análise de interação de grupo, testando para maiores mudanças de volume na dança em comparação com o grupo esportivo. Um aumento significativo na massa cinzenta foi encontrado nas giro precentral (Montreal Neurological Institute (MNI)-coordenadas: x = -16; y = -18; z = 77) e no parahippocampal giro giro (MNI-coordenadas: x = 34; y = -26; z = -20). Os gráficos da caixa mostram as alterações relativas da matéria cinzenta no voxel de pico. * p ≤ 0, 05.
discussão
neste estudo, comparámos os efeitos da participação num programa de dança ou num programa de desporto de fitness físico convencional sobre a função cerebral e o volume em idosos saudáveis. O programa de dança foi uma intervenção recém-desenhada que exigia constantemente a aprendizagem de novas coreografias de dança. O programa esportivo convencional se concentrou principalmente em exercícios motorizados repetitivos. Como descoberta principal, observamos que após 6 meses de treinamento, os volumes no giro pré-Central esquerdo dos dançarinos tinham aumentado mais do que os do grupo esportivo. Após mais 12 meses de treinamento, um aumento adicional de volume foi observado no giro parahippocampal direito dos dançarinos. Os níveis de BDNF aumentaram durante os primeiros 6 meses de treino de dança e voltaram aos valores pré-tratamento após 18 meses. No grupo do esporte convencional, um aumento semelhante na BDNF não era evidente. Como os níveis de aptidão cardiovascular ao longo das intervenções permaneceram constantes em ambos os grupos, os efeitos observados não podiam ser atribuídos a melhorias na aptidão física, mas pareciam estar relacionados com as características específicas do programa de dança. Estas características incluíam a exigência de aprender constantemente novas coreografias( ou seja, memória), integrar informação multisensorial, coordenar todo o corpo e navegar no espaço.
alterações cerebrais
o giro pré-central é essencial para o controlo das funções motoras voluntárias. O aumento do volume de matéria cinzenta no giro pré-central no grupo de dança pode, portanto, ter sido baseado nos padrões de movimento complexos e sempre em mudança que os dançarinos tiveram que realizar. Estes movimentos exigiram a coordenação simultânea de várias partes do corpo em diferentes direções e ajuste aos diversos ritmos da música (policêntrica e polirrítmica). Reflectindo sobre estes complexos requisitos de coordenação, Brown et al. (2006) have reported dance-induced activations in the putamen, the primary motor cortex and the supplementary motor area (SMA), as shown by PET. Outros estudos indicaram uma associação entre as exigências de coordenação (por exemplo, equilíbrio, malabarismo) e neuroplasticidade na região pré-Central (Boyke et al., 2008; Taubert et al., 2010). Assim, o aumento do volume relacionado com a dança nesta área foi consistente com as expectativas baseadas na literatura.
O giro parahippocampal faz parte do arco exterior do sistema límbico e desempenha um papel importante na memória de trabalho e recuperação de memória episódica (Pantel et al., 2003). De acordo com o Êxtase e de Lomo (1973), o parahippocampal giro constitui a interface entre a memória e a experiência da consciência do presente, porque ele está interligado pela perforant trato tanto para as regiões do lobo frontal, que são associadas com a memória de trabalho, e o hipocampo, a estrutura central na memória episódica de codificação e espacial de navegação. Muitos estudos de VBM relataram uma perda de volume relacionada com a idade em regiões parahippocampais (Tisserand et al., 2002). Além disso, Echávarri et al. (2011)sugeriram que a atrofia parahippocampal é um biomarcador precoce do anúncio. A observação de que a realização de um programa de dança por um período mais longo pode induzir processos neuroplásticos nesta região crucial da memória, portanto, particularmente encorajadora em termos de desenvolvimento de estratégias de prevenção.
dinâmica Temporal da plasticidade cerebral da matéria cinzenta
o volume observado aumenta nas duas regiões cerebrais desenvolvidas em momentos diferentes. A dança levou a um aumento de volume nas áreas motoras após 6 meses (Ver também Rehfeld et al., 2016), que permaneceu estável durante os 12 meses seguintes. O aumento de volume associado à dança no giro parahippocampal surgiu mais tarde e foi observado apenas nos dados de 18 meses. A diferente dinâmica temporal na evolução das duas regiões cerebrais pode estar relacionada às diferenças nos mecanismos celulares subjacentes. A pesquisa Animal sugeriu que a angiogênese e a formação de novos dendritos (Thomas et al., 2012) ocorrem rapidamente, enquanto que as alterações no neuropil ocorrem muito mais lentamente (Black et al., 1990). Em humanos, foram observados aumentos rápidos do volume de matéria cinzenta nas regiões pré-frontal após apenas 2 semanas de aprendizagem motora em adultos mais jovens (Taubert et al., 2010). Para induzir neuroplasticidade no hipocampo, foram recomendados períodos de formação mais longos (Erickson et al., 2012; Niemann et al., 2014).curiosamente, os aumentos de volume acima mencionados do nosso estudo estão em contraste com os resultados de Hänggi et al. (2010), que revelou que dançarinos de balé profissionais diminuíram os volumes de matéria cinzenta no córtex pré-motora esquerdo, SMA, Putamen e giro frontal superior e os resultados de Hüfner et al. (2011), que relatam volumes reduzidos em várias regiões do cérebro, incluindo a formação hipocampal anterior em dançarinos profissionais e slackliners em comparação com não-profissionais. No entanto, estes estudos foram estudos observacionais transversais que compararam os cérebros de dançarinos profissionais com os de não-profissionais. Além disso, o nosso grupo de dança consistia em noviços idosos. Tem sido demonstrado que aprender de uma nova habilidade primeiro leva ao recrutamento de recursos neurais adicionais. Mais tarde, quando a habilidade se torna mais automática, menos recursos neuronais são necessários, o que pode levar a reduções de volume em aqueles com experiência de longo prazo. Com base nestas condições iniciais (noviços, adultos mais velhos) e no nosso programa especial de treino de dança, que requeria uma aprendizagem constantemente nova dos padrões de movimento, é possível que os efeitos da especialização induzidos volume diminuam como relatado por Hänggi et al. (2010) não são observados.pensa-se geralmente que a formação motora induz inicialmente o aumento do volume cerebral. No entanto, o treinamento prolongado leva à automatização, que pode ter os efeitos opostos no volume cortical, porque menos controle cortical é necessário após as habilidades motoras terem sido totalmente estabelecidas (Taube, 2008). Nosso Programa de treinamento de dança, portanto, foi especialmente projetado para evitar tal automatização, o que pode explicar por que, pelo menos dentro de 18 meses, não foram observadas reduções de volume cortical em nosso estudo.as funções cognitivas também mostraram desenvolvimento não linear, em que a memória verbal aumentou apenas durante o segundo período de treino. Em relação ao desempenho de atenção, melhorias significativas foram observáveis após apenas 6 meses em ambos os grupos (Rehfeld et al., 2016). Estes resultados corroboram relatórios anteriores sobre os efeitos benéficos das intervenções físicas nos testes neuropsicológicos (Bamadis et al., 2014). No entanto, em dados de capacidade cognitiva, em contraste com os dados do cérebro, não surgiram diferenças de grupo. Outros relataram efeitos superiores da formação cognitiva e física combinada em oposição a intervenções individuais (Oswald et al., 2006). Alargaremos ainda mais as nossas intervenções para testar se as diferenças de grupo na cognição poderão surgir ainda mais tarde.embora VBM seja uma modalidade de imagem que revela mudanças de volume no cérebro, esta técnica não permite conclusões causais sobre os processos neurofisiológicos subjacentes. Neurogênese, sinaptogênese e angiogênese são apenas alguns dos mecanismos que têm sido sugeridos para ser a base das mudanças de volume cerebral (Zatorre et al., 2012). Como mediadores dos efeitos da aptidão cardiovascular no cérebro, fatores de crescimento como BDNF, fator de crescimento semelhante a insulina (IGF) e fator de crescimento nervoso (NGF) estão sendo estudados (Kirk-Sanchez e McGough, 2014). No entanto, em nosso estudo, em contraste com os anteriores (Erickson et al., 2011; Maass et al., 2015), não houve diferenças na aptidão cardiovascular entre os grupos, e os níveis de aptidão não se alteraram durante as intervenções. A última observação foi provavelmente relacionada ao nosso controle da frequência cardíaca individual, que visamos manter na zona aeróbica. No entanto, as alterações do BDNF também foram associadas à actividade física, interacção social e stress positivo (Mattson, 2008), e nem todos os estudos observaram um aumento do BDNF após formação cardiovascular (Vital et al., 2014). Finalmente, a pesquisa em animais sugeriu que a coordenação, mas não o treino de resistência, induz a sinaptogénese e alterações gliais (Black et al., 1990). Juntos, os fatores adicionais que conduzem a secreção de BDNF podem ter sido mais cruciais durante a dança do que durante as atividades de fitness, explicando assim por que apenas os dançarinos mostraram um aumento BDNF nos primeiros 6 meses. A observação de que os níveis de BDNF voltaram aos valores basais nos 12 meses seguintes, enquanto os aumentos de volume foram observados simultaneamente no giro parahippocampal, contudo, indica que deve haver outros factores envolvidos na plasticidade cerebral adulta do que os representados pelos níveis de BDNF no sangue periférico.no que diz respeito aos mecanismos neurobiológicos de plasticidade induzida pelo exercício, também o conceito de reserva cerebral (Satz et al., 2011) deve ser considerada. O conceito de reserva cerebral descreve diferenças individuais em uma neuroplasticidade adaptativa de base aumentada, que fornecem maior capacidade dinâmica para remodelar circuitos corticais para diferentes estressores (Barulli e Stern, 2013; Freret et al., 2015).
perspectivas
os resultados do nosso estudo sugerem que uma intervenção dançante a longo prazo pode ser superior ao exercício físico repetitivo na indução de neuroplasticidade no cérebro humano envelhecido. Presumimos que esta vantagem está relacionada com a natureza multimodal da dança, que combina desafios físicos, cognitivos e coordenativos. Para o nosso conhecimento, este é o primeiro estudo longitudinal e aleatório para recomendar programas de dança como um meio de prevenir a matéria cinzenta e o declínio cognitivo nos idosos. Mais pesquisas são necessárias para esclarecer em maior detalhe a dinâmica temporal e os mecanismos neurobiológicos subjacentes da neuroplasticidade induzida pela dança e se esta intervenção realmente tem o potencial de reduzir o risco de doenças neurodegenerativas como a doença de Alzheimer.
contribuições do autor
PM projetou e realizou a pesquisa, analisou os dados, escreveu o artigo. KR projetou e realizou a pesquisa. A em analisou os dados. AH e VL desenharam a pesquisa. MD: recolha de dados, revisão de artigos. TB e JK projetaram a pesquisa, analisaram os dados. NGM desenhou a pesquisa, escreveu o artigo.
Declaração de conflito de interesses
os autores declaram que a investigação foi realizada na ausência de quaisquer relações comerciais ou financeiras que possam ser interpretadas como um potencial conflito de interesses.Ashburner, J., and Friston, K. J. (2000). Morfometria baseada em Voxel-os métodos. Neuroimagem 11, 805-821. doi: 10.1006 / nimg.2000.0582
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Bamadis, P. D., Vivas, A. B., Styliadis, C., Frantzidis, M., Klados, M., Schlee, W., et al. (2014). A review of physical and cognitive interventions in aging. Neuroci. Biobehav. Rev. 44, 206-220. doi: 10.1016 / j. neubiorev.2014.03.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Barulli, D., and Stern, Y. (2013). Eficiência, capacidade, compensação, manutenção, plasticidade: conceitos emergentes na reserva cognitiva. Trends Cogn. Ciência. 17, 502–509. doi: 10.1016 / j. tics.2013.08.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Black, J. E., Isaacs, K. R., Anderson, B. J., Alcantara, A. A., and Greenough, W. T. (1990). A aprendizagem causa sinaptogénese, enquanto a actividade motora causa angiogénese, no córtex cerebelar de ratos adultos. Procedimento. Natl. Acad. Ciência. U S A 87, 5568-5572. doi: 10.1073 / pnas.87.14.5568
PubMed Abstract | CrossRef Full Text/Google Scholar
Bliss, T. V., and Lomo, T. (1973). Potenciação duradoura da transmissão sináptica no dentado do coelho anaetizado após estimulação da via perforante. J. Physiol. 232, 331–356. doi: 10.1113 / jphysiol.1973.sp010273
PubMed Abstract | CrossRef Full Text/Google Scholar
Boyke, J., Driemeyer, J., Gaser, C., Büchel, C., and May, A. (2008). Alterações da estrutura cerebral induzidas pela formação nos idosos. J. Neurosci. 28, 7031–7035. doi: 10.1523/JNEUROSCI.0742-08.2008
PubMed Abstract | CrossRef Full Text/Google Scholar
Brown, S., Martinez, M., and Parsons, M. (2006). A base neuronal da dança humana. Cérebro. Córtex 16, 1157-1167. doi: 10.1093/cercor/bhj057
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Colcombe, S. J. Erickson, K. I., Scalf, E. P. E., Kim, J. S. Prakash, R., McAuley, E., et al. (2006). O treinamento aeróbico aumenta o volume do cérebro em humanos idosos. J. Gerontol. A. Biol. Ciência. Med. Ciência. 61, 1166–1170. doi: 10.1093 / gerona / 61.11.1166
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Echávarri, C., Aalten, P., Uyling, H. B. M., Jacobs, H. I. L., Visser, P. J., Gronenschild, M., et al. (2011). Atrofia no giro parahippocampal como um biomarcador precoce da doença de Alzheimer. Estrutura Cerebral. Funct. 215, 265–271. doi: 10.1007/s00429-010-0283-8
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Edelmann, E., Lessmann, V., e Brigadski, T. (2014). Torção pré e pós-sináptica na secreção de BDNF e acção na plasticidade sináptica. Neuropharmacology 76, 610-627. doi: 10.1016 / j. neuropharm.2013.05.043
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., et al. (2011). O treinamento de exercício aumenta o tamanho do hipocampo e melhora a memória. Procedimento. Natl. Acad. Ciência. U S A 108, 3017-3022. doi: 10.1073 / pnas.1015950108
PubMed Abstract | CrossRef Full Text | Google Scholar
Erickson, K. I., Weinstein, A. M., and Lopez, O. L. (2012). Actividade física, plasticidade cerebral e doença de Alzheimer. Arco. Med. Res. 43, 615-621. doi: 10.1016 / j. arcmed.2012.09.008
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Flöel, A., Ruscheweyh, R., Krüger, K., Willemer, C., Inverno, B., Völker, K., et al. (2010). Actividade física e funções de memória: as neurotrofinas e o volume de matéria cinzenta cerebral são o elo que falta? Neuroimage 49, 2756-2763. doi: 10.1016 / j. neuroimage.2009.10.043
PubMed Abstract | CrossRef Full Text | Google Scholar
Freret, T., Gaudreau, P., Schumann-Bard, P., Billard, J. M., and Popa-Wagner, A. (2015). Mecanismos subjacentes ao efeito neuroprotector da reserva cerebral contra a depressão do fim da vida. J. Neural. Transm. (Vienna) 122, S55–S61. doi: 10.1007 / s00702-013-1154-2
PubMed Abstract | CrossRef Full Text/Google Scholar
Gregory, S. M., Parker, B., and Thompson, P. D. (2012). Atividade física, função cognitiva e saúde cerebral: Qual é o papel do treinamento de exercício na prevenção da demência? Ciência Cerebral. 2, 684–708. doi: 10.3390/brainsci2040684
PubMed Abstract | CrossRef Full Text | Google Scholar
Hänggi, J., Koeneke, S., Bezzola, L. ,and Jäncke, L. (2010). Neuroplasticidade estrutural na rede sensorimotor de bailarinas profissionais. Cantarolar. MAPP Cerebral. 31, 1196–1206. doi: 10.1002 / hbm.20928
PubMed Abstract | CrossRef Full Text/Google Scholar
Helmstaedter, C., and Durwen, H. F. (1990). O teste de aprendizagem verbal e retenção. Uma ferramenta útil e diferenciada na avaliação do desempenho da memória verbal. Suico. Arco. Neurol. Psychiatr. (1985) 141, 21–30.
PubMed Abstract | Google Scholar
Huang, E. J., and Reichardt, L. F. (2001). Neurotrofinas: Papeis in neuronal development and function. Annu. Rev. Neurosci. 24, 677–736. doi: 10.1146 / annurev.neuro.24.1.677
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Hüfner, K., Binetti, C., Hamilton, D. A., Stephan, T., Flanagin, V. L., Linn, J., et al. (2011). Plasticidade estrutural e funcional da formação hippocampal em dançarinos profissionais e slackliners. Hippocampus 21, 855-865. doi: 10.1002/hipo.20801 PubMed Abstract | CrossRef Full Text/Google Scholar
Karvonen, M. J., Kentala, E., and Mustala, O. (1957). The effects of training on heart rate: a longitudinal study. Anao. Med. Expo. Biol. Fenn. 35, 307–315.
PubMed Abstract/Google Scholar
Kattenstroth, J. C., Kalisch, T., Holt, S., Tegethoff, M., and Dinse, H. R. (2013). Seis meses de intervenção da dança aumenta o desempenho postural, sensorimotor e cognitivo nos idosos sem afetar as funções cardio-respiratórias. Frente. Neurocise Envelhecida. 5:5. doi: 10.3389 / fnagi.2013.00005
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Kattenstroth, J. C., Kolankowska, I., Kalisch, T., and Dinse, H. R. (2010). Desempenho sensorial, motor e cognitivo Superior em indivíduos idosos com atividades de dança multi-ano. Frente. Neurocise Envelhecida. 2:31. doi: 10.3389 / fnagi.2010.00031
PubMed Abstract | CrossRef Full Text | Google Scholar
Kempermann, G., Kuhn, H. G., and Gage, F. H. (1997). Mais neurónios hipocampos em ratos adultos a viver num ambiente enriquecido. Natureza. 386, 493–495. doi: 10.1038/386493a0
PubMed Abstract | CrossRef Full Text | Google Scholar
Kirk-Sanchez, N. J., and McGough, E. L. (2014). Exercício físico e desempenho cognitivo nos idosos: perspectivas actuais. Clin. Interv. Aging 9, 51-62. doi: 10.2147 / CIA.S39506
PubMed Abstract | CrossRef Full Text | Google Scholar
Lessmann, V., and Brigadski, T. (2009). Mechanisms, locations, and kinetics of synaptic BDNF secreção: an update. Neuroci. Res. 65, 11-22. D. O. I.: 10.1016 / j. neures.2009.06.004
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Maass, A., Düzel, S., Goerke, M., Becke, A., Sobieray, U., Neumann, K., et al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M., Hökelmann, A., Kaufmann, J., et al. (2016). Efeitos do treino de dança e desporto de saúde na massa cinzenta do cérebro de idosos saudáveis. Sportwiss. 46, 213-222. doi: 10.1007/s12662-016-0411-6
CrossRef Texto Completo / Google Scholar
Niemann, C., Godde, B., e Voelcker-Rehage, C. (2014). Não só cardiovascular, mas também exercício coordenado aumenta o volume hipocampo em adultos mais velhos. Frente. Neurocise Envelhecida. 6:170. doi: 10.3389 / fnagi.2014.00170
PubMed Abstract / CrossRef Full Text/Google Scholar
Oswald, W. D., Gunzelmann, T., Rupprecht, R., and Hagen, B. (2006). Efeitos diferenciais da formação cognitiva e física única versus combinada com adultos mais velhos: o estudo SimA numa perspectiva de 5 anos. Euro. J. Ageing 3, 179-192. doi: 10.1007 / s10433-006-0035-Z
CrossRef Full Text | Google Scholar
Pantel, J., Kratz, B., Essig, M., and Schröde, J. (2003). Défices do volume Parahippocampal em indivíduos com declínio cognitivo associado ao envelhecimento. Manha. J. Psychiatry 160, 379-382. doi: 10.1176/appi.ajp.160.2.379
PubMed Abstract / CrossRef Full Text | Google Scholar
Park, H., and Poo, M. M. (2013). Regulação neurotrofina do desenvolvimento e função do circuito neural. Conversao. Rev. Neurosci. 14, 7–23. doi: 10.1038/nrn3379
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Rehfeld, K., Hökelmann, A., Kaufmann, J., e Müller, N. G. (2016). “Effects of a dance vs. a fitness training on brain plasticity, balance performance and attention in healthy seniores: a new approach with SPM 12 for pairwise longitudinal group comparison,” in Proceeding-International Conference of Sports and Neuroscience (Magdeburg), 127-138.
Satz, P., Cole, M. A., Hardy, D. J., and Rassovsky, Y. (2011). Cérebro e reserva cognitiva: mediador (s) e validade da construção, uma crítica. J. Clin. Expo. Neuropsicol. 33, 121–130. doi: 10.1080/13803395.2010.493151
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Schega, L., Pedro, B., Brigadski, T., Leßmann, V., Isermann, B., Hamacher, D., et al. (2016). Efeito da hipoxia normobárica intermitente na capacidade aeróbica e na função cognitiva em pessoas idosas. J. Sci. Med. Desporto 19, 941-945. doi: 10.1016 / j. jsams.2016.02.012
PubMed Abstract | CrossRef Full Text/Google Scholar
Taube, W. (2008). Adaptações neurofisiológicas em resposta ao treino de equilíbrio. Dtsch. Z. Sportmed. 63, 273–277. doi: 10.5960 / dzsm.2012.030
CrossRef Texto Completo | Google Scholar
Taubert, M., Draganski, B., Anwander, A., Muller, K., Horstmann, A., Villringer, A., et al. (2010). Propriedades dinâmicas da estrutura cerebral humana: mudanças relacionadas com a aprendizagem em áreas corticais e Conexões de fibra associadas. J. Neurosci. 30, 11670–11677. doi: 10.1523/JNEUROSCI.2567-10.
PubMed Abstract | CrossRef Full Text/Google Scholar
Thomas, A. G., Dennis, A., Bandettini, P. A., and Johansen-Berg, H. (2012). Os efeitos da actividade aeróbica na estrutura cerebral. Frente. Psychol. 3:86. doi: 10.3389/fpsyg.2012.00086
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Tisserand, D. J., Pruessner, J. C., Sanz Arigita, E. J., van Boxtel, M. P., Fonseca, A. C., Jolles, J., et al. (2002). Os volumes corticais frontais regionais diminuem diferentemente no envelhecimento: um estudo de IRM para comparar abordagens volumétricas e morfometria baseada em voxel. Neuroimagem 17, 657-669. doi: 10.1016/s1053-8119(02)91173-0
PubMed Resumo | CrossRef Texto Completo | Google Scholar
van Praag, H., Shubert, T., Zhao, C., e Gage, F. H. (2005). O exercício aumenta a aprendizagem e a neurogenese hipocampal em ratos idosos. J. Neurosci. 25, 8680–8685. doi: 10.1523/JNEUROSCI.1731-05.2005
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Verghese, J., Lipton, R. B., Katz, M. J., Hall, C. B., Dervy, C. A., Kuslansky, G., et al. (2003). Actividades de lazer e risco de demência em idosos. N. Engl. J. Med. 348, 2508–2516. doi: 10.1056/NEJMoa022252
PubMed Resumo | CrossRef Texto Completo | Google Scholar
Vital, T. M.; Stein, A. M., Melo Coelho, F. G., Arantes, F. J., Teodorov, E., Santos-Galduróz, R. F., et al. (2014). Exercício físico e Factor de crescimento endotelial vascular (VEGF) em idosos: uma revisão sistemática. Arco. Gerontol. Geriatr. 59, 234–239. doi: 10.1016 / j. archger.2014.04.011
PubMed Abstract / CrossRef Full Text | Google Scholar
Voelcker-Rehage, C., and Niemann, C. (2013). Alterações cerebrais estruturais e funcionais relacionadas com diferentes tipos de actividade física ao longo da vida. Neuroci. Biobehav. Rev. 37, 2268-2295. doi: 10.1016 / j. neubiorev.2013.01.028
PubMed Abstract / CrossRef Full Text | Google Scholar
Zatorre, R. J., Fields, R. D., and Johansen-Berg, H. (2012). Plasticidade em cinza e branco: alterações neuroimaging na estrutura cerebral durante a aprendizagem. Conversao. Neuroci. 15, 528–536. doi: 10.1038 / nn.3045
PubMed Abstract / CrossRef Full Text/Google Scholar
Zimmermann, P., and Fimm, B. (2002). Test battery for attention testing (TAP), versão 1.7, manual—Part1. Würselen: Psytest.