
Introduzione
Il cambiamento demografico nelle società Occidentali, che coinvolgono sia un parente e un aumento assoluto del numero di persone anziane, ha suscitato un crescente interesse scientifico geriatrica problemi. In questo contesto, i concetti di invecchiamento di successo stanno diventando sempre più importanti. Diversi studi hanno indicato che l’esercizio fisico può svolgere un ruolo chiave nell’invecchiamento sano e nella prevenzione del declino cognitivo e delle malattie neurodegenerative (Colcombe et al., 2006; Erickson et al., 2011). Recensioni recenti che riassumono studi epidemiologici, trasversali e interventistici supportano l’attività fisica come metodo propizio per indurre la neuroplasticità nella tarda età adulta (Gregory et al., 2012; Voelcker-Rehage e Niemann, 2013; Bamadis et al., 2014). Erickson et al. (2012) hanno concluso che una maggiore forma fisica cardiorespiratoria e livelli di attività fisica aerobica sono associati a maggiori volumi di materia grigia nelle regioni prefrontali e nell’ippocampo. Tuttavia, non solo fitness cardiovascolare ma anche esercizio coordinativo (Niemann et al., 2014) e formazione cognitiva (Bamadis et al., 2014) hanno dimostrato di indurre plasticità della materia grigia e migliorare le funzioni cognitive negli adulti più anziani.
La ricerca sugli animali ha suggerito che una combinazione di esercizio fisico con arricchimento sensoriale ha l’effetto più forte sulla genesi di nuovi neuroni—prevalentemente nell’ippocampo—e che solo questa combinazione garantisce la sopravvivenza duratura delle cellule appena nate (Kempermann et al., 1997; van Praag et al., 2005). Kattenstroth et al. (2013) hanno suggerito che “le attività di danza dovrebbero essere considerate come un equivalente di condizioni ambientali arricchite per gli esseri umani poiché forniscono a un individuo maggiori esigenze sensoriali, motorie e cognitive.”Nonostante questa affermazione incoraggiante, gli studi che esaminano gli effetti dell’allenamento di danza sulla struttura o sulla funzione del cervello sono scarsi. Dopo un intervento di danza di 6 mesi, Kattenstroth et al. (2010) hanno riportato miglioramenti significativi nelle prestazioni cognitive, tattili e motorie negli anziani partecipanti. I risultati di uno studio prospettico su 469 soggetti di età superiore ai 75 anni su un periodo di follow-up mediano di 5,1 anni hanno indicato che la danza è associata a un rischio marcatamente ridotto di demenza (Verghese et al., 2003). Tuttavia, Hüfner et al. (2011) hanno riportato volumi ridotti in diverse regioni del cervello tra cui la formazione dell’ippocampo anteriore e parti della corteccia vestibolare parieto-insulare in ballerini professionisti e slackliner rispetto ai non professionisti. Questa scoperta può indicare che l’allenamento intensivo e ripetitivo delle stesse capacità motorie porta nel contesto della specializzazione a volumi ridotti di alcune regioni del cervello. In questo studio, progettiamo quindi uno speciale programma di formazione di danza che richiede costantemente ai partecipanti di apprendere nuovi modelli di movimento (Müller et al., 2016). Per valutare i benefici specifici di questo intervento, abbiamo confrontato il nostro programma di danza di nuova concezione con un gruppo di controllo attivo piuttosto che passivo, che ha preso parte a un convenzionale programma di fitness health sport in cui i partecipanti eseguivano tipicamente esercizi fisici ripetitivi come andare in bicicletta su un ergometro. Inoltre, poiché eravamo interessati alla dinamica temporale degli interventi, abbiamo valutato gli effetti sulla struttura e sulla funzione del cervello dopo 6 e 18 mesi di allenamento. In tal modo, abbiamo cercato di valutare se è utile estendere gli interventi, perché alcune regioni del cervello possono richiedere più formazione di altre, o se c’è un limite dopo il quale più formazione diventa dannosa. Infine, alla ricerca di un potenziale meccanismo alla base della neuroplasticità, abbiamo misurato i livelli di BDNF nel sangue periferico. Diversi studi hanno suggerito che BDNF promuove la differenziazione di nuovi neuroni e sinapsi (Huang e Reichardt, 2001; Lessmann e Brigadski, 2009; Park e Poo, 2013; Edelmann et al., 2014). BDNF, quindi, è stato proposto come mediatore della neuroplasticità adulta (Flöel et al., 2010).
Materiali e metodi
Partecipanti e Progettazione sperimentale
Lo studio è stato progettato come un intervento controllato di 18 mesi. Lo studio è stato approvato dal comitato etico dell’Università Otto-von-Guericke, Magdeburgo, e tutti i soggetti hanno firmato un comunicato scritto e non hanno ricevuto il pagamento per la loro partecipazione. Sessantadue individui anziani sani (63-80 anni) reclutati tramite annunci sui giornali locali sono stati sottoposti a screening per lo studio. I criteri di esclusione erano claustrofobia, tinnito, protesi metalliche, tatuaggi, diabete mellito, depressione (Beck-Depressions Inventory, BDI-II > 13), deficit cognitivi (Mini-Mental State Examination, MMSE < 27), malattie neurologiche ed esercizio fisico regolare (≥1 ora/settimana). Sulla base di questi criteri, sono stati esclusi 10 soggetti. I restanti 52 partecipanti sono stati poi assegnati in modo casuale sia al gruppo di danza o lo sport utilizzando il sito web www.randomization.com e controllo per età, stato MMSE e forma fisica. Le valutazioni sono state eseguite al basale, dopo 6 e dopo 18 mesi di allenamento (Figura 1). Ventidue partecipanti hanno completato l’intero intervento e tutte le misurazioni. La tabella 1 fornisce dati demografici dettagliati per questi partecipanti. Non sono state riscontrate differenze di gruppo per quanto riguarda i dati demografici.
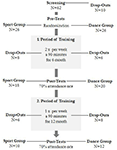
Figura 1. Diagramma di flusso del reclutamento dei partecipanti.
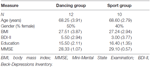
Tabella 1. Informazioni demografiche sui partecipanti al basale.
Interventi
Gli interventi sono stati suddivisi in due periodi. Nel primo periodo, i soggetti si sono allenati due volte a settimana in sessioni di 90 minuti per 6 mesi. Per ragioni pratiche (disponibilità di partecipanti e formatori), il secondo periodo di intervento di 12 mesi ha comportato una frequenza di formazione ridotta di una volta alla settimana. Entrambi i programmi di intervento sono stati eseguiti in un contesto di gruppo con musica per controllare le interazioni psicosociali. Il carico condizionale è stato esaminato registrando i valori degli impulsi durante le sessioni di allenamento e calcolando la frequenza cardiaca di allenamento individuale secondo Karvonen et al. (1957) con un fattore 0.6 per un allenamento di resistenza esteso. Quindi, i due programmi di allenamento erano comparabili in termini di intensità, durata e frequenza. Entrambi sono stati supervisionati da istruttori esperti.
Gruppo di danza
I partecipanti al gruppo di danza hanno partecipato a un programma di formazione di nuova concezione in cui veniva costantemente chiesto loro di apprendere nuove sequenze di movimento. Queste coreografie richiedevano il coordinamento di diverse parti del corpo (cioè gambe, braccia, tronco) nello spazio in diverse condizioni di sforzo (sforzo fisico, precisione, situazione e pressione temporale). I soggetti dovevano imparare le coreografie a memoria, imponendo così elevate esigenze anche alla memoria. Il programma comprendeva cinque generi diversi (line dance, jazz dance, rock “n” roll e square dance), che sono stati cambiati dopo ogni quarta sessione. Nel corso dell’intervento, le richieste coordinative e la pressione temporale sono state aumentate introducendo movimenti di danza e coreografie più complesse e aumentando i battiti al minuto nella musica.
Gruppo sportivo
I partecipanti al gruppo sportivo hanno completato un programma di allenamento forza-resistenza convenzionale con esercizi principalmente ripetitivi e basse richieste in termini di coordinazione e memoria di tutto il corpo. Ogni sessione comprendeva unità di 20 minuti di allenamento di resistenza, resistenza alla forza e flessibilità. L’allenamento di resistenza è stato eseguito su cicloergometri. Nell’unità forza-resistenza sono stati eseguiti movimenti alternati (ad esempio, riccioli bicipiti, squat, sit-up), ma sono stati evitati movimenti complessi di tutto il corpo per mantenere basse le richieste coordinative. L’unità di flessibilità consisteva principalmente in esercizi di stretching.
Outcome Measures
Cardiovascular Fitness
L’idoneità cardiovascolare è stata valutata mediante il test Physical Working Capacity 130 (PWC 130). PWC 130 è la potenza (misurata in watt) che un soggetto è in grado di raggiungere su un cicloergometro con una frequenza cardiaca di 130 bpm. Per i calcoli, abbiamo usato la frequenza cardiaca a riposo e la relativa capacità fisica .
Test neuropsicologici
Una vasta batteria di test neuropsicologici è stata eseguita sui soggetti. Ai fini dello studio attuale, sono riportati solo i risultati di un test verbale di memoria a breve e lungo termine, il “VLMT” (una versione tedesca adattata del “Rey Uditive Verbal Learning Test”; Helmstaedter e Durwen, 1990) e una batteria di test di attenzione (Test di prestazioni attentive (TAP); Zimmermann e Fimm, 2002).
BDNF
Campioni di sangue a digiuno sono stati prelevati al mattino delle valutazioni neuropsicologiche. Dai campioni di sangue, le concentrazioni plasmatiche di BDNF sono state determinate da sandwich ELISAs (BDNF DuoSets; R & D Systems, Wiesbaden, Germania) come precedentemente descritto (Schega et al., 2016).
MRI
Le immagini MR sono state acquisite su un 3 Tesla Siemens MAGNETOM Verio (Syngo MR B17) utilizzando una bobina di testa a 32 canali. Le sequenze di MPRAGE ponderate in T1 ad alta risoluzione sono state acquisite utilizzando un protocollo di imaging 3D rapid gradient echo imaging preparato con magnetizzazione (224 sezioni sagittali, dimensione voxel: 0,8 × 0,8 × 0,8 mm3, TR: 2500 ms, TE: 3,47 ms, TI: 1100 ms, angolo di vibrazione: 7°). Le immagini MR sono state analizzate utilizzando la morfometria basata su voxel (VBM) implementata in SPM 12 (Welcome Department of Cognitive Neurology, Londra, Regno Unito). VBM è una tecnica imparziale a tutto cervello per l’analisi del volume regionale della materia grigia e dei cambiamenti tissutali (Ashburner e Friston, 2000).
La preelaborazione ha coinvolto la segmentazione della materia grigia, la creazione di modelli tramite DARTEL, la normalizzazione spaziale allo spazio standardizzato del Montreal Neurological Institute (MNI) e la levigatura con un kernel gaussiano di 8 mm a larghezza completa a metà massima (FWHM).
Per analizzare la differenza nelle variazioni di volume della materia grigia tra i gruppi, è stato applicato un disegno a fattoriale completo con il gruppo di fattori (danza, sport) e il tempo (0, 6 e 18 mesi). Nel caso di interazioni significative di gruppo × tempo, test t post hoc tra coppie consecutive di punti temporali (0 vs. 6; 0 vs. 18; 6 vs. 18 mesi) sono stati calcolati separatamente per ciascun gruppo. Per tutte le analisi è stata applicata una soglia di p < 0,001 (non corretta).
Risultati
La presentazione dei risultati è strutturata come segue. Abbiamo prima cercato un effetto di intervento generale (fattore tempo), poi abbiamo cercato gli effetti differenziali dei due interventi (gruppo di interazione × tempo); infine, un’analisi più dettagliata della dinamica temporale è stata eseguita tramite confronti a coppie post hoc.
L’idoneità cardiovascolare
L’idoneità cardiovascolare misurata dal PWC 130 (Tabella 2, capacità fisica relativa) non differiva tra i gruppi al basale. Inoltre, il punteggio PWC 130 non è aumentato in modo significativo nel corso del tempo di entrambi gli interventi.

Tabella 2. Mezzi e (SD) per il fitness, il funzionamento cognitivo, i livelli plasmatici di BDNF e il volume totale di materia grigia all’interno dei gruppi di allenamento durante l’intervento.
Test Neuropsicologici
Un significativo effetto principale di tempo è stato osservato per il verbale di memoria a breve termine (VLMT primi di richiamo; F(2,19) = 6.438, p = 0.004, η2 = 0.253), verbale a lungo termine di richiamo libero (VLMT fine di richiamo; F(2,19) = 3.387, p = 0.049, η2 = 0.244), verbale a lungo termine di riconoscimento (VLMT di riconoscimento; F(2,19) = 5.352, p = 0.009, η2 = 0.220) e attenzione ai tempi di reazione (sottoprova flessibilità; F(2,19) = 19.156, p < 0.001, η2 = 0.489). Nessun tempo significativo × interazioni di gruppo emerse.
I confronti a coppie post hoc hanno mostrato miglioramenti significativi dal basale a 18 mesi e da 6 a 18 mesi in tutte e tre le sottocategorie VLMT in entrambi i gruppi. Per quanto riguarda i tempi di reazione TAP, è stato osservato un miglioramento significativo nel confronto tra il basale e i dati a 18 mesi in entrambi i gruppi.
BDNF
I livelli plasmatici di BDNF sono stati analizzati in campioni di sangue prima dell’inizio dell’allenamento e 6 mesi e 18 mesi dopo l’inizio dell’allenamento. I livelli plasmatici assoluti di BDNF sono riassunti nella Tabella 2. I cambiamenti intraindividuali nel livello di BDNF hanno rivelato un aumento significativo nel gruppo di danza, mentre il livello intraindividuale di BDNF è rimasto costante dopo 6 mesi di allenamento nel gruppo sportivo. Non si sono verificate ulteriori variazioni di BDNF dopo 18 mesi in nessun gruppo (Figura 2).
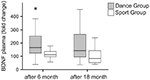
Figura 2. Cambiamenti intraindividuali nei livelli plasmatici di BDNF dopo l’intervento. I livelli plasmatici di BDNF sono stati analizzati in campioni di sangue di partecipanti che eseguono un programma di allenamento di danza o un programma di allenamento sportivo prima dell’inizio dell’allenamento, dopo 6 mesi di allenamento e dopo 18 mesi di allenamento. L’aumento relativo dei livelli di BDNF è stato quantificato. I livelli di BDNF sono aumentati significativamente nel gruppo di danza dopo 6 mesi di allenamento (Mann-Whitney U-test, p < 0.004) e sono diminuiti quasi al valore basale del valore di pretrattamento dopo 18 mesi di allenamento di danza. Non c’è stato alcun cambiamento nell’intero corso del tempo nel gruppo sportivo (test Friedman, p = 0.319), mentre un cambiamento significativo nell’intero corso del tempo è stato osservato nel gruppo di danza (test Friedman, p = 0.028). Box plot: minimo, 25 ° percentile, mediana, 75 ° percentile. * p ≤ 0,05.
MRI
È stata osservata una significativa interazione di gruppo × tempo nel giro precentrale sinistro e nel parahippocampale destro. I test t post hoc tra la linea di base e i dati di 6 mesi hanno mostrato un aumento significativo del volume di materia grigia nel giro precentrale sinistro dei soli ballerini. Nel confronto tra la linea di base e i dati di 18 mesi oltre al giro precentrale, i ballerini hanno mostrato un significativo aumento del volume della materia grigia nel giro parahippocampale destro, che era anche l’unico cambiamento significativo nell’intervallo da 6 a 18 mesi. Pertanto, l’aumento del volume nel giro precentrale è emerso dopo 6 mesi ed è rimasto stabile nel restante intervallo di allenamento della danza, mentre il cambiamento nel giro parahippocampale si è verificato solo durante l’intervallo di allenamento successivo (Figura 3).
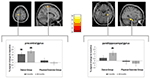
Figura 3. Tempo per analisi di interazione di gruppo, test per maggiori variazioni di volume nella danza rispetto al gruppo sportivo. Un aumento significativo della materia grigia è stato trovato nel giro precentrale (Montreal Neurological Institute (MNI)-coordinate: x = -16; y = -18; z = 77) e nel giro del giro parahippocampale (MNI-coordinate: x = 34; y = -26; z = -20). I grafici a riquadri mostrano le variazioni relative della materia grigia nel voxel di picco. * p ≤ 0,05.
Discussione
In questo studio, abbiamo confrontato gli effetti della partecipazione a un programma di danza oa un programma di sport di forma fisica convenzionale sulla funzione cerebrale e sul volume negli anziani sani. Il programma di danza era un intervento di nuova concezione che richiedeva costantemente l’apprendimento di nuove coreografie di danza. Il programma di sport convenzionale si è concentrato principalmente su esercizi motori ripetitivi. Come risultato principale, abbiamo osservato che dopo 6 mesi di allenamento, i volumi nel giro precentrale sinistro dei ballerini erano aumentati di più rispetto a quelli del gruppo sportivo. Dopo altri 12 mesi di allenamento, è stato osservato un ulteriore aumento del volume nel giro parahippocampale destro dei ballerini. I livelli di BDNF sono aumentati durante i primi 6 mesi di allenamento di danza e sono tornati ai valori pre-trattamento dopo 18 mesi. Nel gruppo sportivo convenzionale, un aumento simile di BDNF non era evidente. Poiché i livelli di fitness cardiovascolare nel corso degli interventi sono rimasti costanti in entrambi i gruppi, gli effetti osservati non potevano essere attribuiti a miglioramenti nella forma fisica, ma sembravano essere correlati alle caratteristiche specifiche del programma di danza. Queste caratteristiche includevano l’esigenza di apprendere costantemente nuove coreografie (cioè la memoria), di integrare informazioni multisensoriali, di coordinare tutto il corpo e di navigare nello spazio.
Cambiamenti cerebrali
Il giro precentrale è essenziale per il controllo delle funzioni motorie volontarie. L’aumento del volume della materia grigia nel giro precentrale nel gruppo di danza potrebbe, quindi, essere stato basato sui complessi e mutevoli schemi di movimento che i ballerini dovevano eseguire. Questi movimenti richiedevano la coordinazione simultanea di diverse parti del corpo in direzioni diverse e l’adeguamento ai ritmi variabili della musica (policentrico e poliritmico). Riflettendo su questi complessi requisiti di coordinamento, Brown et al. (2006) hanno riportato attivazioni indotte dalla danza nel putamen, nella corteccia motoria primaria e nell’area motoria supplementare (SMA), come mostrato dal PET. Altri studi hanno indicato un’associazione tra esigenze di coordinamento (ad esempio, bilanciamento, giocoleria) e neuroplasticità nella regione precentrale (Boyke et al., 2008; Taubert et al., 2010). Quindi, l’aumento del volume legato alla danza in quest’area era coerente con le aspettative basate sulla letteratura.
Il giro parahippocampale fa parte dell’arco esterno del sistema limbico e svolge un ruolo importante nella memoria di lavoro e nel recupero della memoria episodica (Pantel et al., 2003). Secondo Bliss e Lomo (1973), il giro parahippocampale costituisce l’interfaccia tra la memoria e la coscienza esperienziale del presente, perché è interconnesso dal tratto perforante sia alle regioni del lobo frontale, che sono associate alla memoria di lavoro, sia all’ippocampo, la struttura centrale nella codifica della memoria episodica e nella navigazione spaziale. Molti studi VBM hanno riportato una perdita di volume legata all’età nelle regioni parahippocampali (Tisserand et al., 2002). Inoltre, Echávarri et al. (2011) hanno suggerito che l’atrofia parahippocampale è un biomarcatore precoce di AD. L’osservazione che impegnarsi in un programma di danza per un periodo più lungo può indurre processi neuroplastici in questa regione cruciale della memoria, quindi, particolarmente incoraggiante in termini di sviluppo di strategie di prevenzione.
Dinamica temporale della plasticità cerebrale della materia grigia
Il volume osservato aumenta nelle due regioni cerebrali sviluppate in tempi diversi. La danza ha portato ad un aumento del volume nelle aree motorie dopo 6 mesi (vedi anche Rehfeld et al., 2016), che è rimasto stabile durante i successivi 12 mesi. L’aumento del volume associato alla danza nel giro parahippocampale è emerso più tardi ed è stato osservato solo nei dati di 18 mesi. Le diverse dinamiche temporali nell’evoluzione delle due regioni cerebrali possono essere correlate a differenze nei meccanismi cellulari sottostanti. La ricerca sugli animali ha suggerito che l’angiogenesi e la formazione di nuovi dendriti (Thomas et al., 2012) si verificano rapidamente, mentre i cambiamenti nel neuropil si verificano molto più lentamente (Black et al., 1990). Negli esseri umani, sono stati osservati rapidi aumenti del volume di materia grigia nelle regioni prefrontali dopo solo 2 settimane di apprendimento motorio negli adulti più giovani (Taubert et al., 2010). Per indurre neuroplasticità nell’ippocampo, sono stati raccomandati periodi di allenamento più lunghi (Erickson et al., 2012; Niemann et al., 2014).
È interessante notare che i suddetti aumenti di volume del nostro studio sono in contrasto con i risultati di Hänggi et al. (2010), che ha rivelato che i ballerini professionisti hanno diminuito i volumi di materia grigia nella corteccia premotoria sinistra, SMA, Putamen e giro frontale superiore e i risultati di Hüfner et al. (2011), che riportano volumi ridotti in diverse regioni del cervello tra cui la formazione dell’ippocampo anteriore in ballerini professionisti e slackliner rispetto ai non professionisti. Tuttavia, questi studi erano studi osservazionali trasversali che confrontavano il cervello dei ballerini professionisti con quelli dei non professionisti. Inoltre, il nostro gruppo di danza era composto da vecchi novizi invecchiati. È stato dimostrato che l’apprendimento di una nuova abilità porta prima al reclutamento di risorse neurali aggiuntive. Più tardi, quando l’abilità diventa più automatica, sono necessarie meno risorse neuronali che possono portare a diminuzioni di volume in quelli con esperienza a lungo termine. Sulla base di queste condizioni iniziali (novizi, anziani) e del nostro speciale programma di allenamento di danza progettato, che richiedeva un apprendimento costantemente nuovo dei modelli di movimento, è possibile che gli effetti della specializzazione indotta diminuiscano come riportato da Hänggi et al. (2010) non sono stati osservati.
Generalmente si pensa che l’allenamento motorio inizialmente induca aumenti del volume cerebrale. Tuttavia, l’allenamento prolungato porta all’automatizzazione, che può avere gli effetti opposti sul volume corticale, perché è necessario un controllo meno corticale dopo che le capacità motorie sono state completamente stabilite (Taube, 2008). Il nostro programma di allenamento di danza, quindi, è stato appositamente progettato per evitare tale automatizzazione, il che potrebbe spiegare perché, almeno entro 18 mesi, non sono state osservate diminuzioni del volume corticale nel nostro studio.
Le funzioni cognitive hanno anche mostrato uno sviluppo non lineare, per cui la memoria verbale è aumentata solo durante il secondo periodo di allenamento. Per quanto riguarda le prestazioni di attenzione, miglioramenti significativi sono stati osservabili dopo soli 6 mesi in entrambi i gruppi (Rehfeld et al., 2016). Questi risultati supportano relazioni precedenti riguardanti gli effetti benefici degli interventi fisici sui test neuropsicologici (Bamadis et al., 2014). Tuttavia, nei dati sulle capacità cognitive, in contrasto con i dati del cervello, non sono emerse differenze di gruppo. Altri hanno riportato effetti superiori dell’allenamento cognitivo e fisico combinato rispetto ai singoli interventi (Oswald et al., 2006). Estenderemo ulteriormente i nostri interventi per verificare se le differenze di gruppo nella cognizione potrebbero emergere anche in momenti successivi.
Meccanismi cellulari e molecolari sottostanti della plasticità della materia grigia
Sebbene VBM sia una modalità di imaging che rivela cambiamenti di volume nel cervello, questa tecnica non consente conclusioni causali riguardanti i processi neurofisiologici sottostanti. Neurogenesi, sinaptogenesi e angiogenesi sono solo alcuni dei meccanismi che sono stati suggeriti come la base dei cambiamenti del volume cerebrale (Zatorre et al., 2012). Come mediatori degli effetti del fitness cardiovascolare sul cervello, sono allo studio fattori di crescita come BDNF, fattore di crescita simile all’insulina (IGF) e fattore di crescita del nervo (NGF) (Kirk-Sanchez e McGough, 2014). Tuttavia, nel nostro studio, in contrasto con quelli precedenti (Erickson et al., 2011; Maass et al., 2015), non erano presenti differenze nella forma fisica cardiovascolare tra i gruppi e i livelli di fitness non cambiavano durante gli interventi. Quest’ultima osservazione era probabilmente correlata al nostro controllo della frequenza cardiaca individuale, che miravamo a mantenere nella zona aerobica. Tuttavia, i cambiamenti di BDNF sono stati associati anche all’attività fisica, all’interazione sociale e allo stress positivo (Mattson, 2008), e non tutti gli studi hanno osservato un aumento di BDNF dopo l’allenamento cardiovascolare (Vital et al., 2014). Infine, la ricerca sugli animali ha suggerito che il coordinamento, ma non l’allenamento di resistenza, induce sinaptogenesi e cambiamenti gliali (Black et al., 1990). Insieme, i fattori aggiuntivi nominati che guidano la secrezione di BDNF potrebbero essere stati più cruciali durante la danza che durante le attività di fitness, spiegando così perché solo i ballerini hanno mostrato un aumento di BDNF nei primi 6 mesi. L’osservazione che i livelli di BDNF sono tornati al basale nei successivi 12 mesi mentre gli aumenti di volume sono stati osservati simultaneamente nel giro parahippocampale, tuttavia, indica che ci devono essere altri fattori coinvolti nella plasticità del cervello adulto rispetto a quelli rappresentati dai livelli di BDNF nel sangue periferico.
Per quanto riguarda i meccanismi neurobiologici della plasticità indotta dall’esercizio, anche il concetto di riserva cerebrale (Satz et al., 2011) dovrebbe essere considerato. Il concetto di riserva cerebrale descrive le differenze individuali in una maggiore neuroplasticità adattiva al basale, che fornisce una maggiore capacità dinamica per il rimodellamento dei circuiti corticali a diversi fattori di stress (Barulli e Stern, 2013; Freret et al., 2015).
Prospettive
I risultati del nostro studio suggeriscono che un intervento di danza a lungo termine potrebbe essere superiore all’esercizio fisico ripetitivo nell’indurre la neuroplasticità nell’invecchiamento del cervello umano. Presumiamo che questo vantaggio sia legato alla natura multimodale della danza, che combina sfide fisiche, cognitive e coordinative. A nostra conoscenza, questo è il primo studio longitudinale randomizzato a raccomandare programmi di danza come mezzo per prevenire la materia grigia e il declino cognitivo negli anziani. Sono necessarie ulteriori ricerche per chiarire in modo più dettagliato le dinamiche temporali e i meccanismi neurobiologici sottostanti alla neuroplasticità indotta dalla danza e se questo intervento abbia veramente il potenziale per ridurre il rischio di malattie neurodegenerative come l’Alzheimer.
Contributi dell’autore
PM ha progettato ed eseguito la ricerca, analizzato i dati, scritto l’articolo. KR ha progettato ed eseguito la ricerca. MS ha analizzato i dati. AH e VL hanno progettato la ricerca. MD: raccolta dati, revisione articolo. TB e JK hanno progettato la ricerca, analizzato i dati. NGM ha progettato la ricerca, ha scritto l’articolo.
Dichiarazione sul conflitto di interessi
Gli autori dichiarano che la ricerca è stata condotta in assenza di relazioni commerciali o finanziarie che potrebbero essere interpretate come un potenziale conflitto di interessi.
Ashburner, J., e Friston, KJ (2000). Morfometria basata su Voxel-i metodi. Neuroimage 11, 805-821. doi: 10.1006 / nimg.2000.0582
PubMed Abstract | CrossRef Full Text/Google Scholar
Bamadis, PD, Vivas, AB, Styliadis, C., Frantzidis, M., Klados, M., Schlee, W., et al. (2014). Una revisione degli interventi fisici e cognitivi nell’invecchiamento. Neurosci. Biobehav. Apoc 44, 206-220. doi: 10.1016 / j. neubiorev.2014.03.019
PubMed Abstract | CrossRef Full Text/Google Scholar
Barulli, D., and Stern, Y. (2013). Efficienza, capacità, compensazione, manutenzione, plasticità: concetti emergenti nella riserva cognitice. Tendenze Cogn. Sic. 17, 502–509. doi: 10.1016 / j. tic.2013.08.012
PubMed Abstract | CrossRef Full Text/Google Scholar
Nero, J. E., Isaacs, K. R., Anderson, B. J., Alcantara, A. A., e Greenough, W. T. (1990). L’apprendimento causa la sinaptogenesi, mentre l’attività motoria causa l’angiogenesi, nella corteccia cerebellare dei ratti adulti. Proc. Natl. Acad. Sic. U S A 87, 5568-5572. doi: 10.1073 / pnas.87.14.5568
PubMed Abstract | CrossRef Full Text/Google Scholar
Bliss, TV, e Lomo, T. (1973). Potenziamento di lunga durata della trasmissione sinaptica nel dentato del coniglio anestetizzato a seguito della stimolazione del percorso perforante. J. Physiol. 232, 331–356. doi: 10.1113 / jphysiol.1973.sp010273
PubMed Abstract | CrossRef Full Text/Google Scholar
Boyke, J., Driemeyer, J., Gaser, C., Büchel, C. e May, A. (2008). Cambiamenti della struttura cerebrale indotti dall’allenamento negli anziani. J. Neurosci. 28, 7031–7035. doi: 10.1523 / JNEUROSCI.0742-08.2008
PubMed Abstract | CrossRef Full Text/Google Scholar
Brown, S., Martinez, M., and Parsons, M. (2006). La base neuronale della danza umana. Cereb. Cortex 16, 1157-1167. doi: 10.1093/cercor/bhj057
PubMed Abstract / CrossRef Full Text/Google Scholar
Colcombe, S. J., Erickson, K. I., Scalf, PE, Kim, J. S., Prakash, R., McAuley, E., et al. (2006). L’allenamento aerobico aumenta il volume del cervello negli esseri umani che invecchiano. J. Gerontol. A. Biol. Sic. Med. Sic. 61, 1166–1170. doi: 10.1093/gerona / 61.11.1166
PubMed Abstract | CrossRef Full Text/Google Scholar
Echávarri, C., Aalten, P., Uyling, H. B. M., Jacobs, H. I. L., Visser, P. J., Gronenschild, M., et al. (2011). Atrofia nel giro parahippocampale come biomarcatore precoce della malattia di Alzheimer. Struttura cerebrale. Funzione. 215, 265–271. doi: 10.1007 / s00429-010-0283-8
PubMed Abstract | CrossRef Full Text/Google Scholar
Edelmann, E., Lessmann, V. e Brigadski, T. (2014). Torsioni pre e postsinaptiche nella secrezione di BDNF e azione nella plasticità sinaptica. Neurofarmacologia 76, 610-627. doi: 10.1016 / j.neuropharm.2013.05.043
PubMed Abstract | CrossRef Full Text/Google Scholar
Erickson, KI, Voss, MW, Prakash, RS, Basak, C., Szabo, A., Chaddock, L., et al. (2011). L’allenamento fisico aumenta le dimensioni dell’ippocampo e migliora la memoria. Proc. Natl. Acad. Sic. U S A 108, 3017-3022. doi: 10.1073 / pnas.1015950108
PubMed Abstract | CrossRef Full Text/Google Scholar
Erickson, KI, Weinstein, A. M. e Lopez, O. L. (2012). Attività fisica, plasticità cerebrale e malattia di Alzheimer. Arco. Med. Res. 43, 615-621. doi: 10.1016 / j.arcmed.2012.09.008
PubMed Abstract | CrossRef Full Text/Google Scholar
Flöel, A., Rusceweyh, R., Krüger, K., Willemer, C., Winter, B., Völker, K., et al. (2010). Attività fisica e funzioni di memoria: le neurotrofine e il volume della materia grigia cerebrale sono l’anello mancante? Neuroimage 49, 2756-2763. doi: 10.1016 / j.neuroimage.2009.10.043
PubMed Abstract | CrossRef Full Text/Google Scholar
Freret, T., Gaudreau, P., Schumann-Bard, P., Billard, JM, e Popa-Wagner, A. (2015). Meccanismi alla base dell’effetto neuroprotettivo della riserva cerebrale contro la depressione tardiva. J. Neurale. Transm. (Vienna) 122, S55-S61. doi: 10.1007 / s00702-013-1154-2
PubMed Abstract | CrossRef Full Text/Google Scholar
Gregory, S. M., Parker, B. e Thompson, PD (2012). Attività fisica, funzione cognitiva e salute del cervello: qual è il ruolo dell’allenamento nella prevenzione della demenza? Cervello Sci. 2, 684–708. doi: 10.3390/brainsci2040684
PubMed Abstract / CrossRef Full Text/Google Scholar
Hänggi, J., Koeneke, S., Bezzola, L., e Jäncke, L. (2010). Neuroplasticità strutturale nella rete sensomotoria di ballerine professionali. Ronzio. Cervello Mapp. 31, 1196–1206. doi: 10.1002 / hbm.20928
PubMed Abstract | CrossRef Full Text/Google Scholar
Helmstaedter, C., and Durwen, HF (1990). Il test di apprendimento e ritenzione verbale. Uno strumento utile e differenziato nella valutazione delle prestazioni della memoria verbale. Svizzera. Arco. Neurolo. Psichiatria. (1985) 141, 21–30.
PubMed Abstract/Google Scholar
Huang, EJ, e Reichardt, LF (2001). Neurotrofine: ruoli nello sviluppo e nella funzione neuronale. Annu. Rev. Neurosci. 24, 677–736. doi: 10.1146 / annurev.neuro.24.1.677
PubMed Abstract | CrossRef Full Text/Google Scholar
Hüfner, K., Binetti, C., Hamilton, DA, Stephan, T., Flanagin, VL, Linn, J., et al. (2011). Plasticità strutturale e funzionale della formazione dell’ippocampo in ballerini professionisti e slackliners. Ippocampo 21, 855-865. doi: 10.1002 / hipo.20801
PubMed Abstract / CrossRef Full Text/Google Scholar
Karvonen, MJ, Kentala, E., e Mustala, O. (1957). Gli effetti dell’allenamento sulla frequenza cardiaca: uno studio longitudinale. Ann. Med. Scad. Biol. Fenn. 35, 307–315.
PubMed Abstract/Google Scholar
Kattenstroth, JC, Kalisch, T., Holt, S., Tegethoff, M., e Dinse, HR (2013). Sei mesi di intervento di danza migliorano le prestazioni posturali, sensomotorie e cognitive negli anziani senza influire sulle funzioni cardio-respiratorie. Anteriore. Invecchiamento Neurosci. 5:5. doi: 10.3389 / fnagi.2013.00005
PubMed Abstract | CrossRef Full Text/Google Scholar
Kattenstroth, JC, Kolankowska, I., Kalisch, T., e Dinse, HR (2010). Prestazioni sensoriali, motorie e cognitive superiori in individui anziani con attività di danza pluriennali. Anteriore. Invecchiamento Neurosci. 2:31. doi: 10.3389 / fnagi.2010.00031
PubMed Abstract | CrossRef Full Text/Google Scholar
Kempermann, G., Kuhn, HG, and Gage, FH (1997). Più neuroni ippocampali nei topi adulti che vivono in un ambiente arricchito. Natura. 386, 493–495. doi: 10.1038/386493a0
PubMed Abstract | CrossRef Full Text/Google Scholar
Kirk-Sanchez, NJ, e McGough, EL (2014). Esercizio fisico e prestazioni cognitive negli anziani: prospettive attuali. Clin. Interv. Invecchiamento 9, 51-62. doi: 10.2147 / CIA.S39506
PubMed Abstract | CrossRef Full Text/Google Scholar
Lessmann, V., e Brigadski, T. (2009). Meccanismi, posizioni e cinetica della secrezione sinaptica di BDNF: un aggiornamento. Neurosci. Res. 65, 11-22. doi: 10.1016 / j. neures.2009.06.004
PubMed Abstract | CrossRef Full Text/Google Scholar
Maass, A., Düzel, S., Goerke, M., Becke, A., Sobieray, U., Neumann, K., et al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M., Hökelmann, A., Kaufmann, J., et al. (2016). Effetti dell’allenamento sportivo di danza e salute sulla materia grigia del cervello degli anziani sani. Sciupafemmine. 46, 213-222. doi: 10.1007 / s12662-016-0411-6
CrossRef Full Text/Google Scholar
Niemann, C., Godde, B. e Voelcker-Rehage, C. (2014). Non solo l’esercizio cardiovascolare, ma anche coordinato aumenta il volume dell’ippocampo negli adulti più anziani. Anteriore. Invecchiamento Neurosci. 6:170. doi: 10.3389 / fnagi.2014.00170
PubMed Abstract / CrossRef Full Text / Google Scholar
Oswald, WD, Gunzelmann, T., Rupprecht, R., and Hagen, B. (2006). Effetti differenziali dell’allenamento cognitivo e fisico singolo rispetto a quello combinato con gli adulti più anziani: lo studio SimA in una prospettiva di 5 anni. EUR. J. Invecchiamento 3, 179-192. doi: 10.1007 / s10433-006-0035-z
CrossRef Full Text/Google Scholar
Pantel, J., Kratz, B., Essig, M., e Schröde, J. (2003). Deficit di volume parahippocampale in soggetti con declino cognitivo associato all’invecchiamento. Essere. J. Psichiatria 160, 379-382. doi: 10.1176 / appi.aip.160.2.379
PubMed Abstract / CrossRef Full Text / Google Scholar
Park, H., e Poo, M. M. (2013). Regolazione delle neurotrofine dello sviluppo e della funzione dei circuiti neurali. NAT. Rev. Neurosci. 14, 7–23. doi: 10.1038 | nrn3379
PubMed Abstract/CrossRef Full Text/Google Scholar
Rehfeld, K., Hökelmann, A., Kaufmann, J., and Müller, NG (2016). “Effetti di una danza contro un allenamento di fitness sulla plasticità del cervello, sull’equilibrio delle prestazioni e sull’attenzione nei seniores sani: un nuovo approccio con SPM 12 per il confronto longitudinale a coppie”, in Prosieguo-Conferenza internazionale di sport e neuroscienze (Magdeburgo), 127-138.
Satz, P., Cole, M. A., Hardy, DJ, e Rassovsky, Y. (2011). Brain and cognitive reserve: mediator (s) and construct validity, a critique. J. Clin. Scad. Neuropsicolo. 33, 121–130. doi: 10.1080 | 13803395.2010.493151
PubMed Abstract/CrossRef Full Text/Google Scholar
Schega, L., Peter, B., Brigadski, T., Leßmann, V., Isermann, B., Hamacher, D., et al. (2016). Effetto dell’ipossia normobarica intermittente sulla capacità aerobica e sulla funzione cognitiva nelle persone anziane. J. Sic. Med. Sport 19, 941-945. doi: 10.1016 / j. jsams.2016.02.012
PubMed Abstract | CrossRef Full Text/Google Scholar
Taube, W. (2008). Adattamenti neurofisiologici in risposta all’allenamento dell’equilibrio. Dtsch. Z. Sportmed. 63, 273–277. doi: 10.5960 / dzsm.2012.030
CrossRef Full Text/Google Scholar
Taubert, M., Draganski, B., Anwander, A., Müller, K., Horstmann, A., Villringer, A., et al. (2010). Proprietà dinamiche della struttura del cervello umano: cambiamenti correlati all’apprendimento nelle aree corticali e connessioni delle fibre associate. J. Neurosci. 30, 11670–11677. doi: 10.1523 / JNEUROSCI.2567-10.2010
PubMed Abstract | CrossRef Full Text/Google Scholar
Thomas, A. G., Dennis, A., Bandettini, P. A., e Johansen-Berg, H. (2012). Gli effetti dell’attività aerobica sulla struttura del cervello. Anteriore. Psicol. 3:86. doi: 10.3389 / fpsyg.2012.00086
PubMed Abstract | CrossRef Full Text/Google Scholar
Tisserand, DJ, Pruessner, JC, Sanz Arigita, EJ, van Boxtel, MP, Evans, AC, Jolles, J., et al. (2002). I volumi corticali frontali regionali diminuiscono in modo differenziato nell’invecchiamento: uno studio MRI per confrontare approcci volumetrici e morfometria basata su voxel. Neuroimage 17, 657-669. doi: 10.1016/s1053-8119(02)91173-0
PubMed Abstract | CrossRef Full Text/Google Scholar
van Praag, H., Shubert, T., Zhao, C., e Gage, FH (2005). L’esercizio migliora l’apprendimento e la neurogenesi ippocampale nei topi invecchiati. J. Neurosci. 25, 8680–8685. doi: 10.1523 / JNEUROSCI.1731-05.2005
PubMed Abstract | CrossRef Full Text | Google Scholar
Verghese, J., Lipton, R. B., Katz, M. J., Hall, C. B., Dervy, C. A., Kuslansky, G., et al. (2003). Attività ricreative e il rischio di demenza negli anziani. N. Ingl. J. Med. 348, 2508–2516. doi: 10.1056/NEJMoa022252
PubMed Abstract / CrossRef Full Text/Google Scholar
Vital, T. M., Stein, A. M., de Melo Coelho, F. G., Arantes, F. J., Teodorov, E., Santos-Galduróz, R. F., et al. (2014). Esercizio fisico e fattore di crescita endoteliale vascolare (VEGF) negli anziani: una revisione sistematica. Arco. Gerontol. Geriatr. 59, 234–239. doi: 10.1016 / j. archger.2014.04.011
PubMed Abstract / CrossRef Full Text / Google Scholar
Voelcker-Rehage, C., e Niemann, C. (2013). Cambiamenti strutturali e funzionali del cervello legati a diversi tipi di attività fisica durante l’arco della vita. Neurosci. Biobehav. Apoc 37, 2268-2295. doi: 10.1016 / j. neubiorev.2013.01.028
PubMed Abstract | CrossRef Full Text/Google Scholar
Zatorre, RJ, Fields, RD, and Johansen-Berg, H. (2012). Plasticità in grigio e bianco: cambiamenti di neuroimaging nella struttura del cervello durante l’apprendimento. NAT. Neurosci. 15, 528–536. doi: 10.1038 / nn.3045
PubMed Abstract / CrossRef Full Text/Google Scholar
Zimmermann, P., and Fimm, B. (2002). Batteria di prova per il test di attenzione (TAP), versione 1.7, manuale—Part1. Würselen: Psytest.