
Einleitung
Der gegenwärtige demografische Wandel in den westlichen Gesellschaften, der sowohl einen relativen als auch einen absoluten Anstieg der Zahl älterer Menschen mit sich bringt, hat ein zunehmendes wissenschaftliches Interesse an geriatrischen Fragen geweckt. In diesem Zusammenhang gewinnen Konzepte des erfolgreichen Alterns zunehmend an Bedeutung. Mehrere Studien haben gezeigt, dass körperliche Bewegung eine Schlüsselrolle beim gesunden Altern und bei der Prävention von kognitivem Verfall und neurodegenerativen Erkrankungen spielen kann (Colcombe et al., 2006; Erickson et al., 2011). Jüngste Reviews, die epidemiologische, Querschnitts- und interventionelle Studien zusammenfassen, unterstützen körperliche Aktivität als günstige Methode zur Induzierung von Neuroplastizität im späten Erwachsenenalter (Gregory et al., 2012; Voelcker-Rehage und Niemann, 2013; Bamadis et al., 2014). Erickson et al. (2012) sind zu dem Schluss gekommen, dass eine höhere kardiorespiratorische Fitness und aerobe körperliche Aktivität mit einem größeren Volumen der grauen Substanz in den präfrontalen Regionen und im Hippocampus verbunden sind. Aber nicht nur kardiovaskuläre Fitness, sondern auch koordinatives Training (Niemann et al., 2014) und kognitives Training (Bamadis et al., 2014) haben gezeigt, dass sie die Plastizität der grauen Substanz induzieren und die kognitiven Funktionen bei älteren Erwachsenen verbessern.
Tierversuche legen nahe, dass eine Kombination aus körperlicher Betätigung und sensorischer Anreicherung den stärksten Einfluss auf die Entstehung neuer Neuronen — vorwiegend im Hippocampus — hat und dass nur diese Kombination das dauerhafte Überleben der neugeborenen Zellen sichert (Kempermann et al., 1997; van Praag et al., 2005). Kattenstroth et al. (2013) haben vorgeschlagen, dass „Tanzaktivitäten als Äquivalent zu angereicherten Umweltbedingungen für den Menschen angesehen werden sollten, da sie einem Individuum erhöhte sensorische, motorische und kognitive Anforderungen stellen.“ Trotz dieser ermutigenden Aussage gibt es nur wenige Studien, die die Auswirkungen von Tanztraining auf die Struktur oder Funktion des Gehirns untersuchen. Nach einer 6-monatigen Tanzintervention haben Kattenstroth et al. (2010) berichteten über signifikante Verbesserungen der kognitiven, taktilen und motorischen Leistung bei teilnehmenden Senioren. Die Ergebnisse einer prospektiven Studie mit 469 Probanden, die älter als 75 Jahre waren, über einen medianen Follow-up-Zeitraum von 5,1 Jahren haben gezeigt, dass Tanzen mit einem deutlich reduzierten Demenzrisiko verbunden ist (Verghese et al., 2003). Hüfner et al. (2011) berichteten über reduzierte Volumina in mehreren Hirnregionen, einschließlich der anterioren Hippocampusbildung und Teilen des parietoinsulären vestibulären Kortex bei professionellen Tänzern und Slacklinern im Vergleich zu Nicht-Profis. Dieser Befund könnte darauf hindeuten, dass intensives und sich wiederholendes Training der gleichen motorischen Fähigkeiten im Kontext der Spezialisierung zu reduzierten Volumina einiger Gehirnregionen führt. In dieser Studie entwerfen wir daher ein spezielles Tanztrainingsprogramm, bei dem die Teilnehmer ständig neue Bewegungsmuster erlernen mussten (Müller et al., 2016). Um die spezifischen Vorteile dieser Intervention zu bewerten, verglichen wir unser neu gestaltetes Tanzprogramm eher mit einer aktiven als mit einer passiven Kontrollgruppe, die an einem herkömmlichen Gesundheitssport-Fitnessprogramm teilnahm, bei dem die Teilnehmer typischerweise sich wiederholende körperliche Übungen wie Radfahren auf einem Ergometer durchführten. Da wir außerdem an der zeitlichen Dynamik der Interventionen interessiert waren, bewerteten wir die Auswirkungen auf die Gehirnstruktur und -funktion nach 6 und 18 Monaten Training. Dabei wollten wir beurteilen, ob es vorteilhaft ist, Interventionen zu verlängern, da einige Gehirnregionen möglicherweise mehr Training erfordern als andere, oder ob es eine Grenze gibt, nach der mehr Training schädlich wird. Auf der Suche nach einem möglichen Mechanismus, der der Neuroplastizität zugrunde liegt, haben wir schließlich die BDNF-Spiegel im peripheren Blut gemessen. Mehrere Studien haben gezeigt, dass BDNF die Differenzierung neuer Neuronen und Synapsen fördert (Huang und Reichardt, 2001; Lessmann und Brigadski, 2009; Park und Poo, 2013; Edelmann et al., 2014). BDNF wurde daher als Mediator der adulten Neuroplastizität vorgeschlagen (Flöel et al., 2010).
Materialien und Methoden
Teilnehmer und experimentelles Design
Die Studie wurde als 18-monatige kontrollierte Intervention konzipiert. Die Studie wurde von der Ethikkommission der Otto-von-Guericke-Universität Magdeburg genehmigt, und alle Probanden unterzeichneten eine schriftliche Erklärung und erhielten keine Zahlung für ihre Teilnahme. Zweiundsechzig gesunde ältere Menschen (63-80 Jahre), die über Ankündigungen in lokalen Zeitungen rekrutiert wurden, wurden für die Studie gescreent. Die Ausschlusskriterien waren Klaustrophobie, Tinnitus, Metallimplantate, Tattoos, Diabetes mellitus, Depression (Beck-Depressions Inventory, BDI-II > 13), kognitive Defizite (Mini-Mental State Examination, MMSE < 27), neurologische Erkrankungen und regelmäßige Bewegung (≥1 h/Woche). Auf der Grundlage dieser Kriterien wurden 10 Probanden ausgeschlossen. Der Rest 52 Die Teilnehmer wurden dann über die Website zufällig entweder der Tanz- oder der Sportgruppe zugewiesen www.randomization.com und Kontrolle für Alter, MMSE-Status und körperliche Fitness. Die Bewertungen wurden zu Studienbeginn, nach 6 und nach 18 Monaten Training durchgeführt (Abbildung 1). Zweiundzwanzig Teilnehmer absolvierten die gesamte Intervention und alle Messungen. Tabelle 1 enthält detaillierte demografische Daten für diese Teilnehmer. Es wurden keine Gruppenunterschiede in Bezug auf die demografischen Daten festgestellt.
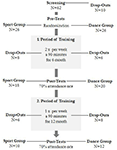
Abbildung 1. Flussdiagramm der Rekrutierung der Teilnehmer.
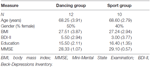
Tabelle 1. Demografische Informationen zu den Teilnehmern zu Studienbeginn.
Interventionen
Die Interventionen wurden in zwei Perioden unterteilt. In der ersten Periode trainierten die Probanden zweimal pro Woche in 90-minütigen Sitzungen für 6 Monate. Aus praktischen Gründen (Verfügbarkeit von Teilnehmern und Trainern) umfasste die zweite 12-monatige Interventionsperiode eine reduzierte Trainingsfrequenz von einmal pro Woche. Beide Interventionsprogramme wurden in einem Gruppenkontext mit Musik zur Kontrolle psychosozialer Interaktionen durchgeführt. Die konditionelle Belastung wurde durch Aufzeichnung der Pulswerte während der Trainingseinheiten und durch Berechnung der individuellen Trainingsherzfrequenz nach Karvonen et al. (1957) mit einem Faktor 0,6 für ausgiebiges Ausdauertraining. Daher waren die beiden Trainingsprogramme in Intensität, Dauer und Häufigkeit vergleichbar. Beide wurden von erfahrenen Instruktoren betreut.
Tanzgruppe
Die Teilnehmer der Tanzgruppe besuchten ein neu gestaltetes Trainingsprogramm, in dem sie ständig aufgefordert wurden, neue Bewegungsabläufe zu erlernen. Diese Choreografien erforderten die Koordination verschiedener Körperteile (d.h. Beine, Arme, Rumpf) im Raum unter verschiedenen Belastungsbedingungen (körperliche Belastung, Präzision, Situation und Zeitdruck). Die Probanden mussten die Choreografien auswendig lernen, was auch hohe Anforderungen an das Gedächtnis stellte. Das Programm umfasste fünf verschiedene Genres (Line Dance, Jazz Dance, Rock „n“ Roll und Square Dance), die nach jeder vierten Session gewechselt wurden. Im Laufe der Intervention wurden koordinative Anforderungen und Zeitdruck erhöht, indem komplexere Tanzbewegungen und Choreografien eingeführt und die Schläge pro Minute in der Musik erhöht wurden.
Sportgruppe
Die Teilnehmer der Sportgruppe absolvierten ein konventionelles Kraft-Ausdauer-Trainingsprogramm mit überwiegend repetitiven Übungen und geringen Anforderungen an die Ganzkörperkoordination und das Gedächtnis. Jede Sitzung umfasste 20-minütige Einheiten Ausdauer-, Kraft-Ausdauer- und Flexibilitätstraining. Das Ausdauertraining wurde an Fahrradergometern durchgeführt. In der Kraft-Ausdauer-Einheit wurden abwechselnde Bewegungen (z. B. Bizeps-Curls, Kniebeugen, Sit-ups) durchgeführt, aber komplexe Ganzkörperbewegungen wurden vermieden, um die koordinativen Anforderungen gering zu halten. Die Flexibilitätseinheit bestand hauptsächlich aus Dehnübungen.
Outcome Measures
Kardiovaskuläre Fitness
Die kardiovaskuläre Fitness wurde mit dem Physical Working Capacity 130 Test (PWC 130) bewertet. PWC 130 ist die Leistung (gemessen in Watt), die ein Proband auf einem Fahrradergometer bei einer Herzfrequenz von 130 Schlägen pro Minute erreichen kann. Für die Berechnungen verwendeten wir die Ruheherzfrequenz und die relative körperliche Kapazität .
Neuropsychologische Tests
An den Probanden wurde eine umfangreiche Batterie neuropsychologischer Tests durchgeführt. Für die Zwecke der aktuellen Studie werden nur die Ergebnisse eines verbalen Kurz- und Langzeitgedächtnistests, des „VLMT“ (eine adaptierte deutsche Version des „Rey Auditory Verbal Learning Test“; Helmstaedter und Durwen, 1990) und einer Aufmerksamkeitstestbatterie (Test of Attentional Performance (TAP); Zimmermann und Fimm, 2002) berichtet.
BDNF
Am Morgen der neuropsychologischen Untersuchungen wurden Nüchternblutproben entnommen. Aus den Blutproben wurden Plasmakonzentrationen von BDNF durch Sandwich-ELISAs (BDNF DuoSets; R&D Systems, Wiesbaden, Deutschland) wie zuvor beschrieben bestimmt (Schega et al., 2016).
MRT
MR-Bilder wurden auf einem 3 Tesla Siemens MAGNETOM Verio (Syngo MR B17) unter Verwendung einer 32-Kanal-Kopfspule aufgenommen. Hochauflösende T1-gewichtete MPRAGE-Sequenzen wurden unter Verwendung eines 3D-Magnetisierungs-präparierten Rapid Gradient Echo Imaging Protocol (224 Sagittalschnitte, Voxelgröße: 0,8 × 0,8 × 0,8 mm3, TR: 2500 ms, TE: 3,47 ms, TI: 1100 ms, Flip-Winkel: 7 °). Die MR-Bilder wurden mit Voxel-basierter Morphometrie (VBM) analysiert, die in SPM 12 (British Department of Cognitive Neurology, London, UK) implementiert wurde. VBM ist eine unvoreingenommene Ganzhirntechnik zur Analyse des regionalen Volumens der grauen Substanz und der Gewebeveränderungen (Ashburner und Friston, 2000).Die Vorverarbeitung umfasste die Segmentierung der grauen Substanz, die Erstellung von Vorlagen über DARTEL, die räumliche Normalisierung auf den standardisierten Raum des Montreal Neurological Institute (MNI) und die Glättung mit einem Gaußschen Kern von 8 mm voller Breite bei halbem Maximum (FWHM).Um den Unterschied in den Volumenänderungen der grauen Substanz zwischen den Gruppen zu analysieren, wurde ein vollfaktorielles Design mit den Faktoren Gruppe (Tanz, Sport) und Zeit (0, 6 und 18 Monate) angewendet. Bei signifikanten Gruppen- × Zeitinteraktionen Post-hoc-t-Tests zwischen aufeinanderfolgenden Zeitpunktpaaren (0 vs. 6; 0 vs. 18; 6 vs. 18 Monate) wurden für jede Gruppe separat berechnet. Für alle Analysen wurde ein Schwellenwert von p < 0,001 (unkorrigiert) angewendet.
Ergebnisse
Die Darstellung der Ergebnisse ist wie folgt strukturiert. Wir suchten zuerst nach einem allgemeinen Interventionseffekt (Faktorzeit), dann nach differentiellen Effekten der beiden Interventionen (Interaktionsgruppe × Zeit); Schließlich wurde eine detailliertere Analyse der zeitlichen Dynamik über paarweise Post-hoc-Vergleiche durchgeführt.
Kardiovaskuläre Fitness
Die kardiovaskuläre Fitness, gemessen mit dem PWC 130 (Tabelle 2, relative körperliche Leistungsfähigkeit), unterschied sich zu Studienbeginn nicht zwischen den Gruppen. Darüber hinaus stieg der PWC 130-Score im Zeitverlauf keiner der beiden Interventionen signifikant an.

Tabelle 2. Mittelwert und (SD) für Fitness, kognitive Funktion, BDNF-Plasmaspiegel und Gesamtvolumen der grauen Substanz innerhalb von Trainingsgruppen über die Intervention.
Neuropsychologische Tests
Ein signifikanter Haupteffekt der Zeit wurde für das verbale Kurzzeitgedächtnis beobachtet (VLMT early recall; F(2,19) = 6,438, p = 0,004, η2 = 0,253), verbaler langzeitfreier Rückruf (VLMT late recall; F(2,19) = 3,387, p = 0,049, η2 = 0,244), verbaler langzeitfreier Rückruf termerkennung (VLMT-Erkennung; F(2,19) = 5,352, p = 0,009, η2 = 0,220) und Aufmerksamkeitsreaktionszeit (Subtestflexibilität; F(2,19) = 19,156, p < 0,001, η2 = 0,489). Es traten keine signifikanten Zeit- oder Gruppeninteraktionen auf.Post-hoc-paarweise Vergleiche zeigten signifikante Verbesserungen von der Baseline bis 18 Monate und von 6 bis 18 Monate in allen drei VLMT-Unterkategorien in beiden Gruppen. In Bezug auf die TAP-Reaktionszeiten wurde eine signifikante Verbesserung im Vergleich der Baseline zu den 18-Monats-Daten in beiden Gruppen beobachtet.
BDNF
Die Plasmaspiegel von BDNF wurden in Blutproben vor Trainingsbeginn sowie 6 Monate und 18 Monate nach Trainingsbeginn analysiert. Die absoluten BDNF-Plasmaspiegel sind in Tabelle 2 zusammengefasst. Die intraindividuellen Veränderungen des BDNF-Spiegels zeigten einen signifikanten Anstieg in der Tanzgruppe, während der intraindividuelle BDNF-Spiegel nach 6 Monaten Training in der Sportgruppe konstant blieb. Nach 18 Monaten traten in keiner Gruppe weitere BDNF-Veränderungen auf (Abbildung 2).
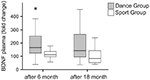
Abbildung 2. Intraindividuelle Veränderungen der BDNF-Plasmaspiegel nach Intervention. BDNF-Plasmaspiegel wurden in Blutproben von Teilnehmern analysiert, die vor Beginn des Trainings, nach 6 Monaten Training und nach 18 Monaten Training ein Tanztrainingsprogramm oder ein Sporttrainingsprogramm durchführten. Der relative Anstieg der BDNF-Spiegel wurde quantifiziert. Die BDNF-Spiegel stiegen in der Tanzgruppe nach 6-monatigem Training signifikant an (Mann-Whitney U-Test, p < 0.004) und sanken nach 18-monatigem Tanztraining fast auf den Ausgangswert des Vorbehandlungswerts. In der Sportgruppe gab es keine Veränderung über den gesamten Zeitverlauf (Friedman-Test, p = 0,319), während in der Tanzgruppe eine signifikante Veränderung über den gesamten Zeitverlauf beobachtet wurde (Friedman-Test, p = 0,028). Box Plots: Minimum, 25. Perzentil, Median, 75. Perzentil. *p ≤ 0,05.
MRT
Im linken Gyrus precentralis und im rechten Parahippocampus wurde eine signifikante Gruppeninteraktion beobachtet. Post-hoc-t-Tests zwischen den Baseline- und 6-Monats-Daten zeigten eine signifikante Zunahme des Volumens der grauen Substanz im linken präzentralen Gyrus nur der Tänzer. Im Vergleich der Baseline- und der 18-Monats-Daten zusätzlich zum präzentralen Gyrus zeigten die Tänzer eine signifikante Volumenzunahme der grauen Substanz im rechten Gyrus Parahippocampus, was auch die einzige signifikante Veränderung im Intervall von 6 bis 18 Monaten war. So trat die Volumenzunahme im Gyrus precentralis nach 6 Monaten auf und blieb über das verbleibende Tanztrainingsintervall stabil, während die Veränderung im Gyrus Parahippocampus nur während des späteren Trainingsintervalls auftrat (Abbildung 3).
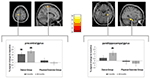
Abbildung 3. Zeit durch Gruppeninteraktionsanalyse, Testen auf größere Volumenänderungen im Tanz im Vergleich zur Sportgruppe. Ein signifikanter Anstieg der grauen Substanz wurde im präzentralen Gyrus (Montreal Neurological Institute (MNI)-Koordinaten: x = -16; y = -18; z = 77) und im Gyrus Parahippocampus Gyrus (MNI-Koordinaten: x = 34; y = -26; z = -20) festgestellt. Die Boxplots zeigen die relativen Graustufenänderungen im Peak-Voxel. *p ≤ 0,05.
Diskussion
In dieser Studie verglichen wir die Auswirkungen der Teilnahme an einem Tanzprogramm oder einem herkömmlichen Fitnesssportprogramm auf die Gehirnfunktion und das Volumen bei gesunden Senioren. Das Tanzprogramm war eine neu gestaltete Intervention, bei der ständig neue Tanzchoreografien gelernt werden mussten. Das konventionelle Sportprogramm konzentrierte sich hauptsächlich auf sich wiederholende motorische Übungen. Als Hauptergebnis beobachteten wir, dass nach 6 Monaten Training die Volumina im linken präzentralen Gyrus der Tänzer stärker zugenommen hatten als in der Sportgruppe. Nach weiteren 12 Monaten Training wurde eine zusätzliche Volumenzunahme im rechten Gyrus Parahippocampus der Tänzer beobachtet. Der BDNF-Spiegel stieg in den ersten 6 Monaten des Tanztrainings an und kehrte nach 18 Monaten zu den Werten vor der Behandlung zurück. In der konventionellen Sportgruppe war ein ähnlicher Anstieg des BDNF nicht offensichtlich. Da die kardiovaskuläre Fitness im Verlauf der Interventionen in beiden Gruppen konstant blieb, konnten die beobachteten Effekte nicht auf Verbesserungen der körperlichen Fitness zurückgeführt werden, sondern schienen mit den spezifischen Merkmalen des Tanzprogramms in Zusammenhang zu stehen. Zu diesen Merkmalen gehörte die Anforderung, ständig neue Choreografien (d. H. Gedächtnis) zu lernen, multisensorische Informationen zu integrieren, den gesamten Körper zu koordinieren und im Raum zu navigieren.
Gehirnveränderungen
Der Gyrus precentralis ist essentiell für die Kontrolle der freiwilligen motorischen Funktionen. Die Zunahme des Volumens der grauen Substanz im präzentralen Gyrus in der Tanzgruppe kann daher auf den komplexen und sich ständig ändernden Bewegungsmustern beruhen, die die Tänzer ausführen mussten. Diese Bewegungen erforderten die gleichzeitige Koordination mehrerer Körperteile in verschiedene Richtungen und die Anpassung an die unterschiedlichen Rhythmen der Musik (polyzentrisch und polyrhythmisch). In Anbetracht dieser komplexen Koordinationsanforderungen haben Brown et al. (2006) berichteten über tanzinduzierte Aktivierungen im Putamen, im primären motorischen Kortex und im ergänzenden motorischen Bereich (SMA), wie von PET gezeigt. Andere Studien haben einen Zusammenhang zwischen Koordinationsanforderungen (z. B. Balancieren, Jonglieren) und Neuroplastizität in der präzentralen Region gezeigt (Boyke et al., 2008; Taubert et al., 2010). Daher entsprach der tanzbedingte Volumenzuwachs in diesem Bereich den Erwartungen, die auf der Literatur beruhten.
Der Gyrus Parahippocampus ist Teil des äußeren Bogens des limbischen Systems und spielt eine wichtige Rolle beim Abrufen des Arbeitsgedächtnisses und des episodischen Gedächtnisses (Pantel et al., 2003). Nach Bliss und Lomo (1973) bildet der Gyrus Parahippocampus die Schnittstelle zwischen dem Gedächtnis und dem Erfahrungsbewusstsein der Gegenwart, da er durch den perforanten Trakt sowohl mit Regionen des Frontallappens, die mit dem Arbeitsgedächtnis assoziiert sind, als auch mit dem Hippocampus, der zentralen Struktur, verbunden ist in der episodischen Gedächtniscodierung und räumlichen Navigation. Viele VBM-Studien haben einen altersbedingten Volumenverlust in parahippocampalen Regionen berichtet (Tisserand et al., 2002). Darüber hinaus Echávarri et al. (2011) haben vorgeschlagen, dass die Parahippocampusatrophie ein früher Biomarker für AD ist. Die Beobachtung, dass die Teilnahme an einem Tanzprogramm über einen längeren Zeitraum neuroplastische Prozesse in dieser entscheidenden Gedächtnisregion auslösen kann, ist daher besonders ermutigend für die Entwicklung von Präventionsstrategien.
Zeitliche Dynamik der Plastizität des Gehirns der grauen Substanz
Die beobachtete Volumenzunahme in den beiden Gehirnregionen entwickelte sich zu unterschiedlichen Zeiten. Tanzen führte nach 6 Monaten zu einer Volumenzunahme der motorischen Areale (siehe auch Rehfeld et al., 2016), die in den folgenden 12 Monaten stabil blieb. Die tanz-assoziierte Volumenzunahme im Gyrus Parahippocampus trat später auf und wurde nur in den 18-Monats-Daten beobachtet. Die unterschiedliche zeitliche Dynamik in der Entwicklung der beiden Hirnregionen kann mit Unterschieden in den zugrunde liegenden zellulären Mechanismen zusammenhängen. Tierversuche haben gezeigt, dass Angiogenese und die Bildung neuer Dendriten (Thomas et al., 2012) treten schnell auf, während Veränderungen im Neuropil viel langsamer auftreten (Black et al., 1990). Beim Menschen wurde nach nur 2 Wochen motorischem Lernen bei jüngeren Erwachsenen ein rascher Anstieg des Volumens der grauen Substanz in den präfrontalen Regionen beobachtet (Taubert et al., 2010). Um die Neuroplastizität im Hippocampus zu induzieren, wurden längere Trainingsperioden empfohlen (Erickson et al., 2012; Niemann et al., 2014).Interessanterweise stehen die oben genannten Volumenzunahmen aus unserer Studie im Gegensatz zu den Ergebnissen von Hänggi et al. (2010), die zeigten, dass professionelle Balletttänzer das Volumen der grauen Substanz im linken prämotorischen Kortex, SMA, Putamen und Gyrus frontalis superior verringert haben, und die Ergebnisse von Hüfner et al. (2011), die über reduzierte Volumina in mehreren Hirnregionen einschließlich der vorderen Hippocampusbildung bei professionellen Tänzern und Slacklinern im Vergleich zu Nicht-Profis berichten. Diese Studien waren jedoch Querschnitt Beobachtungsstudien, die die Gehirne von professionellen Tänzern mit denen von Nicht-Profis verglichen. Darüber hinaus bestand unsere Tanzgruppe aus alten Novizen. Es hat sich gezeigt, dass das Erlernen einer neuen Fähigkeit zunächst zur Rekrutierung zusätzlicher neuronaler Ressourcen führt. Später, wenn die Fertigkeit automatischer wird, werden weniger neuronale Ressourcen benötigt, was bei Personen mit langjähriger Erfahrung zu einer Verringerung des Volumens führen kann. Basierend auf diesen Anfangsbedingungen (Anfänger, ältere Erwachsene) und unserem speziell entwickelten Tanztrainingsprogramm, das ständig neues Lernen von Bewegungsmustern erforderte, ist es möglich, dass die Auswirkungen der Spezialisierung das Volumen verringern, wie von Hänggi et al. (2010) nicht beachtet.
Es wird allgemein angenommen, dass motorisches Training anfänglich eine Zunahme des Gehirnvolumens induziert. Ein längeres Training führt jedoch zu einer Automatisierung, die die gegenteiligen Auswirkungen auf das kortikale Volumen haben kann, da nach vollständiger Etablierung der motorischen Fähigkeiten weniger kortikale Kontrolle erforderlich ist (Taube, 2008). Unser Tanztrainingsprogramm wurde daher speziell entwickelt, um eine solche Automatisierung zu vermeiden, was erklären könnte, warum in unserer Studie zumindest innerhalb von 18 Monaten keine Abnahme des kortikalen Volumens beobachtet wurde.
Kognitive Funktionen zeigten ebenfalls eine nichtlineare Entwicklung, wobei das verbale Gedächtnis erst während der zweiten Trainingsperiode zunahm. In Bezug auf die Aufmerksamkeitsleistung waren in beiden Gruppen bereits nach 6 Monaten signifikante Verbesserungen zu beobachten (Rehfeld et al., 2016). Diese Ergebnisse stützen frühere Berichte über die positiven Auswirkungen physikalischer Interventionen auf neuropsychologische Tests (Bamadis et al., 2014). In den kognitiven Fähigkeitsdaten traten jedoch im Gegensatz zu den Gehirndaten keine Gruppenunterschiede auf. Andere berichteten über überlegene Effekte von kombiniertem kognitivem und körperlichem Training im Gegensatz zu Einzelinterventionen (Oswald et al., 2006). Wir werden unsere Interventionen weiter ausbauen, um zu testen, ob Gruppenunterschiede in der Kognition auch zu späteren Zeitpunkten auftreten können.
Zugrunde liegende zelluläre und molekulare Mechanismen der Plastizität der grauen Substanz
Obwohl VBM eine Bildgebungsmodalität ist, die Volumenänderungen im Gehirn aufdeckt, erlaubt diese Technik keine kausalen Rückschlüsse auf die zugrunde liegenden neurophysiologischen Prozesse. Neurogenese, Synaptogenese und Angiogenese sind nur einige der Mechanismen, die als Grundlage für Hirnvolumenänderungen vorgeschlagen wurden (Zatorre et al., 2012). Als Mediatoren der Auswirkungen der kardiovaskulären Fitness auf das Gehirn werden Wachstumsfaktoren wie BDNF, insulinähnlicher Wachstumsfaktor (IGF) und Nervenwachstumsfaktor (NGF) untersucht (Kirk-Sanchez und McGough, 2014). Doch in unserer Studie, im Gegensatz zu früheren (Erickson et al., 2011; Maass et al., 2015), es gab keine Unterschiede in der kardiovaskulären Fitness zwischen den Gruppen, und das Fitnessniveau änderte sich während der Interventionen nicht. Die letztere Beobachtung hing wahrscheinlich mit unserer Kontrolle der individuellen Herzfrequenz zusammen, die wir in der aeroben Zone aufrechterhalten wollten. BDNF-Veränderungen wurden jedoch auch mit körperlicher Aktivität, sozialer Interaktion und positivem Stress in Verbindung gebracht (Mattson, 2008), und nicht alle Studien haben einen BDNF-Anstieg nach Herz-Kreislauf-Training beobachtet (Vital et al., 2014). Schließlich hat die Tierforschung vorgeschlagen, dass Koordination, aber nicht Ausdauertraining Synaptogenese und Gliaveränderungen induziert (Black et al., 1990). Zusammengenommen könnten die genannten zusätzlichen Faktoren, die die BDNF-Sekretion antreiben, beim Tanzen entscheidender gewesen sein als bei Fitnessaktivitäten, was erklärt, warum nur Tänzer in den ersten 6 Monaten einen BDNF-Anstieg zeigten. Die Beobachtung, dass die BDNF-Spiegel in den folgenden 12 Monaten auf den Ausgangswert zurückkehrten, während gleichzeitig Volumenzunahmen im Gyrus Parahippocampus beobachtet wurden, deutet jedoch darauf hin, dass an der Plastizität des Gehirns bei erwachsenen andere Faktoren beteiligt sein müssen als die BDNF-Spiegel im peripheren Blut.
In Bezug auf neurobiologische Mechanismen der stressinduzierten Plastizität auch Konzept der Gehirnreserve (Satz et al. 2011) berücksichtigt werden. Das Konzept der Gehirnreserve beschreibt individuelle Unterschiede in einer erhöhten adaptiven Neuroplastizität zu Studienbeginn, die eine größere dynamische Kapazität für die Umgestaltung kortikaler Schaltkreise für verschiedene Stressoren bieten (Barulli und Stern, 2013; Freret et al., 2015).
Perspektiven
Die Ergebnisse unserer Studie legen nahe, dass eine langfristige Tanzintervention der repetitiven körperlichen Betätigung bei der Induktion von Neuroplastizität im alternden menschlichen Gehirn überlegen sein könnte. Wir gehen davon aus, dass dieser Vorteil mit der Multimodalität des Tanzens zusammenhängt, die körperliche, kognitive und koordinative Herausforderungen kombiniert. Unseres Wissens ist dies die erste longitudinale, randomisierte Studie, die Tanzprogramme als Mittel zur Verhinderung von grauer Substanz und kognitivem Verfall bei älteren Menschen empfiehlt. Weitere Untersuchungen sind erforderlich, um die zeitliche Dynamik und die zugrunde liegenden neurobiologischen Mechanismen der tanzinduzierten Neuroplastizität genauer zu klären und ob diese Intervention wirklich das Potenzial hat, das Risiko für neurodegenerative Erkrankungen wie Alzheimer zu verringern.
Autorenbeiträge
PM entwarf und führte die Forschung durch, analysierte die Daten und schrieb den Artikel. KR entwarf und führte die Forschung durch. MS analysierte die Daten. AH und VL entwarfen die Forschung. MD: Datenerfassung, Artikelrevision. TB und JK entwarfen die Forschung, analysierten die Daten. NGM entwarf die Forschung, schrieb den Artikel.
Erklärung zum Interessenkonflikt
Die Autoren erklären, dass die Forschung in Abwesenheit von kommerziellen oder finanziellen Beziehungen durchgeführt wurde, die als potenzieller Interessenkonflikt ausgelegt werden könnten.
Ashburner, J., und Friston, KJ (2000). Voxel-basierte Morphometrie-die Methoden. Neuroimage 11, 805-821. doi: 10.1006/nimg.2000.0582
PubMed Zusammenfassung | CrossRef Volltext / Google Scholar
Bamadis, P. D., Vivas, A. B., Styliadis, C., Frantzidis, M., Klados, M., Schlee, W., et al. (2014). Ein Überblick über körperliche und kognitive Interventionen beim Altern. Neurowissenschaften. Biobehav. Offenbarung 44, 206-220. doi: 10.1016/j.neubiorev.2014.03.019
PubMed Abstract / CrossRef Full Text / Google Scholar
Barulli, D. und Stern, Y. (2013). Effizienz, Kapazität, Kompensation, Wartung, Plastizität: aufkommende Konzepte in der kognitiven Reserve. Trends Cogn. Sci. 17, 502–509. doi: 10.1016/j.tics.2013.08.012
PubMed Abstract | CrossRef Full Text / Google Scholar
Black, J. E., Isaacs, K. R., Anderson, B. J., Alcantara, A. A. und Greenough, W. T. (1990). Lernen verursacht Synaptogenese, während motorische Aktivität Angiogenese verursacht, in der Kleinhirnrinde von erwachsenen Ratten. Prok. Natl. Acad. Sci. U S A 87, 5568-5572. doi: 10.1073/pnas.87.14.5568
PubMed Abstract / CrossRef Full Text / Google Scholar
Bliss, T. V. und Lomo, T. (1973). Langanhaltende Potenzierung der synaptischen Transmission im Dentat des anosthetisierten Kaninchens nach Stimulation des perforanten Pfades. In: J. Physiol. 232, 331–356. in: 10.1113/jphysiol.1973.sp010273
PubMed Abstract | CrossRef Full Text / Google Scholar
Boyke, J., Driemeyer, J., Gaser, C., Büchel, C. und May, A. (2008). Trainingsinduzierte Veränderungen der Gehirnstruktur bei älteren Menschen. In: J. Neurosci. 28, 7031–7035. doi: 10.1523/JNEUROSCI.0742-08.2008
PubMed Abstract / CrossRef Full Text / Google Scholar
Brown, S., Martinez, M. und Parsons, M. (2006). Die neuronale Basis des menschlichen Tanzes. In: Cereb. Kapitel 16, 1157-1167. doi: 10.1093/cercor/bhj057
PubMed Zusammenfassung / CrossRef Volltext / Google Scholar
Colcombe, S. J., Erickson, K. I., Scalf, PE, Kim, J. S., Prakash, R., McAuley, E., et al. (2006). Aerobic-Training erhöht das Gehirnvolumen bei alternden Menschen. In: J. Gerontol. IN: Biol. Sci. Med. Sci. 61, 1166–1170. Ursprungsbezeichnung: 10.1093/gerona/61.11.1166
PubMed Abstract | CrossRef Full Text / Google Scholar
Echávarri, C., Aalten, P., Uyling, H. B. M., Jacobs, H. I. L., Visser, P. J., Gronenschild, M., et al. (2011). Atrophie im Gyrus Parahippocampus als früher Biomarker der Alzheimer-Krankheit. Gehirnstruktur. Funct. 215, 265–271. ust-idnr.: 10.1007/s00429-010-0283-8
PubMed Abstract | CrossRef Full Text / Google Scholar
Edelmann, E., Lessmann, V. und Brigadski, T. (2014). Prä- und postsynaptische Wendungen in der BDNF-Sekretion und Wirkung in der synaptischen Plastizität. Neuropharmakologie 76, 610-627. doi: 10.1016/j.neuropharm.2013.05.043
PubMed Zusammenfassung | CrossRef Volltext / Google Scholar
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., et al. (2011). Bewegungstraining erhöht die Größe des Hippocampus und verbessert das Gedächtnis. Prok. Natl. Acad. Sci. U S A 108, 3017-3022. doi: 10.1073/pnas.1015950108
PubMed Abstract / CrossRef Full Text / Google Scholar
Erickson, K. I., Weinstein, A. M. und Lopez, O. L. (2012). Körperliche Aktivität, Plastizität des Gehirns und Alzheimer-Krankheit. Bogen. Med. 43, 615-621. doi: 10.1016/j.arcmed.2012.09.008
PubMed Zusammenfassung | CrossRef Volltext / Google Scholar
Flöel, A., Ruscheweyh, R., Krüger, K., Willemer, C., Winter, B., Völker, K., et al. (2010). Körperliche Aktivität und Gedächtnisfunktionen: Sind Neurotrophine und zerebrale graue Substanz das fehlende Glied? Neuroimage 49, 2756-2763. doi: 10.1016/j.neuroimage.2009.10.043
PubMed Abstract | CrossRef Full Text / Google Scholar
Freret, T., Gaudreau, P., Schumann-Bard, P., Billard, J. M. und Popa-Wagner, A. (2015). Mechanismen, die der neuroprotektiven Wirkung der Gehirnreserve gegen Depressionen im späten Leben zugrunde liegen. J. Neural. In: Transm. (Wien) 122, S55-S61. ust-idnr.: 10.1007/s00702-013-1154-2
PubMed Abstract | CrossRef Full Text / Google Scholar
Gregory, S. M., Parker, B. und Thompson, P. D. (2012). Körperliche Aktivität, kognitive Funktion und Gehirngesundheit: Welche Rolle spielt Bewegungstraining bei der Prävention von Demenz? Gehirn-Sci. 2, 684–708. doi: 10.3390/brainsci2040684
PubMed Zusammenfassung / CrossRef Volltext / Google Scholar
Hänggi, J., Koeneke, S., Bezzola, L., und Jäncke, L. (2010). Strukturelle Neuroplastizität im sensomotorischen Netzwerk professioneller Balletttänzerinnen. Summen. Gehirn Mapp. 31, 1196–1206. doi: 10.1002/hbm.20928
PubMed Abstract | CrossRef Full Text / Google Scholar
Helmstaedter, C. und Durwen, H. F. (1990). Der verbale Lern- und Retentionstest. Ein nützliches und differenziertes Werkzeug zur Bewertung der verbalen Gedächtnisleistung. Schweiz. Bogen. Neurol. Psychiatrie. (1985) 141, 21–30.
PubMed Abstract / Google Scholar
Huang, EJ und Reichardt, LF (2001). Neurotrophine: Rollen in der neuronalen Entwicklung und Funktion. Jahr. Rev. Neurosci. 24, 677–736. doi: 10.1146/Jahrbuch.neuro.24.1.677
PubMed Abstract | CrossRef Full Text / Google Scholar
Hüfner, K., Binetti, C., Hamilton, D. A., Stephan, T., Flanagin, V. L., Linn, J., et al. (2011). Strukturelle und funktionelle Plastizität der Hippocampusformation bei professionellen Tänzern und Slacklinern. Hippocampus 21, 855-865. doi: 10.1002/hipo.20801
PubMed Abstract / CrossRef Full Text / Google Scholar
Karvonen, M. J., Kentala, E., und Mustala, O. (1957). Die Auswirkungen des Trainings auf die Herzfrequenz: eine Längsschnittstudie. Ann. Med. Verwendbar bis. Biol. Fenn. 35, 307–315.
PubMed Abstract / Google Scholar
Kattenstroth, JC, Kalisch, T., Holt, S., Tegethoff, M. und Dinse, HR (2013). Sechs Monate Tanzintervention verbessern die posturale, sensomotorische und kognitive Leistung bei älteren Menschen, ohne die kardiorespiratorischen Funktionen zu beeinträchtigen. Front. In: Aging Neuroscience. 5:5. doi: 10.3389/fnagi.2013.00005
PubMed Zusammenfassung / CrossRef Volltext / Google Scholar
Kattenstroth, J. C., Kolankowska, I., Kalisch, T., und Dinse, HR (2010). Überlegene sensorische, motorische und kognitive Leistung bei älteren Menschen mit mehrjährigen Tanzaktivitäten. Front. In: Aging Neuroscience. 2:31. doi: 10.3389/fnagi.2010.00031
PubMed Abstract | CrossRef Full Text / Google Scholar
Kempermann, G., Kuhn, H. G. und Gage, F. H. (1997). Mehr Hippocampus-Neuronen in erwachsenen Mäusen, die in einer angereicherten Umgebung leben. Natur. 386, 493–495. doi: 10.1038/386493a0
PubMed Zusammenfassung / CrossRef Volltext / Google Scholar
Kirk-Sanchez, NJ, und McGough, EL (2014). Körperliche Bewegung und kognitive Leistungsfähigkeit bei älteren Menschen: aktuelle Perspektiven. Clin. Interv. Altern 9, 51-62. doi: 10.2147/CIA.S39506
PubMed Zusammenfassung / CrossRef Volltext / Google Scholar
Lessmann, V. und Brigadski, T. (2009). Mechanismen, Orte und Kinetik der synaptischen BDNF-Sekretion: ein Update. Neurowissenschaften. 65, 11-22. Ursprungsbezeichnung: 10.1016/j.neures.2009.06.004
PubMed Zusammenfassung | CrossRef Volltext / Google Scholar
Maass, A., Düzel, S., Goerke, M., Becke, A., Sobieray, U., Neumann, K., et al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M., Hökelmann, A., Kaufmann, J., et al. (2016). Effekte eine Tanz- und eines Gesundheitssporttrainings auf die graue Hirnsubstanz gesunder Senioren. Sportwiss. 46, 213–222. doi: 10.1007/s12662-016-0411-6
CrossRef Full Text | Google Scholar
Niemann, C., Godde, B., and Voelcker-Rehage, C. (2014). Not only cardiovascular, but also coordinative exercise increases hippocampal volume in older adults. Front. Aging Neurosci. 6:170. doi: 10.3389/fnagi.2014.00170
PubMed Abstract | CrossRef Full Text | Google Scholar
Oswald, W. D., Gunzelmann, T., Rupprecht, R., und Hagen, B. (2006). Differentielle Effekte von einfachem versus kombiniertem kognitivem und körperlichem Training bei älteren Erwachsenen: Die SimA-Studie in einer 5-Jahres-Perspektive. EUR. J., 3, 179-192. ust-IDNR.: 10.1007/s10433-006-0035- z
CrossRef Full Text / Google Scholar
Pantel, J., Kratz, B., Essig, M. und Schröde, J. (2003). Parahippocampale Volumendefizite bei Probanden mit altersbedingtem kognitivem Verfall. Uhr. J. Psychiatrie 160, 379-382. Ursprungsbezeichnung: 10.1176/appi.ajp.160.2.379
PubMed Zusammenfassung / CrossRef Volltext / Google Scholar
Park, H., und Poo, M. M. (2013). Neurotrophin-Regulation der Entwicklung und Funktion neuronaler Schaltkreise. Nat. Rev. Neurosci. 14, 7–23. doi: 10.1038/nrn3379
PubMed Abstract | CrossRef Full Text / Google Scholar
Rehfeld, K., Hökelmann, A., Kaufmann, J. und Müller, N. G. (2016). „Auswirkungen eines Tanzes vs. eines Fitnesstrainings auf die Plastizität des Gehirns, die Gleichgewichtsleistung und die Aufmerksamkeit bei gesunden Senioren: ein neuer Ansatz mit SPM 12 für den paarweisen longitudinalen Gruppenvergleich“, in Proceeding-International Conference of Sports and Neuroscience (Magdeburg), 127-138.
Satz, P., Cole, M. A., Hardy, D. J. und Rassovsky, Y. (2011). Gehirn und kognitive Reserve: Mediator (e) und Konstruktvalidität, eine Kritik. J. Clin. Verwendbar bis. Neuropsychol. 33, 121–130. doi: 10.1080/13803395.2010.493151
PubMed Zusammenfassung | CrossRef Volltext / Google Scholar
Schega, L., Peter, B., Brigadski, T., Leßmann, V., Isermann, B., Hamacher, D., et al. (2016). Wirkung der intermittierenden normobaren Hypoxie auf die aerobe Kapazität und kognitive Funktion bei älteren Menschen. J. Sci. Med. Sport 19, 941-945. doi: 10.1016/j.jsam.2016.02.012
PubMed Zusammenfassung / CrossRef Volltext / Google Scholar
Taube, W. (2008). Neurophysiologische Anpassungen als Reaktion auf Gleichgewichtstraining. Dtsch. In: Z. Sportmed. 63, 273–277. doi: 10.5960/dzsm.2012.030
CrossRef Volltext / Google Scholar
Taubert, M., Draganski, B., Anwander, A., Müller, K., Horstmann, A., Villringer, A., et al. (2010). Dynamische Eigenschaften der menschlichen Gehirnstruktur: Lernbezogene Veränderungen in kortikalen Bereichen und damit verbundenen Faserverbindungen. In: J. Neurosci. 30, 11670–11677. doi: 10.1523/JNEUROSCI.2567-10.2010
PubMed Abstract / CrossRef Full Text / Google Scholar
Thomas, A. G., Dennis, A., Bandettini, P. A. und Johansen-Berg, H. (2012). Die Auswirkungen der aeroben Aktivität auf die Gehirnstruktur. Front. Psychol. 3:86. doi: 10.3389/fpsyg.2012.00086
PubMed Zusammenfassung | CrossRef Volltext / Google Scholar
Tisserand, D. J., Pruessner, J. C., Sanz Arigita, E. J., van Boxtel, M. P., Evans, A. C., Jolles, J., et al. (2002). Regionale frontale kortikale Volumina nehmen im Alter unterschiedlich ab: eine MRT-Studie zum Vergleich volumetrischer Ansätze und voxelbasierter Morphometrie. Neuroimage 17, 657-669. doi: 10.1016/s1053-8119(02)91173-0
PubMed Zusammenfassung | CrossRef Volltext | Google Scholar
van Praag, H., Shubert, T., Zhao, C. und Gage, F. H. (2005). Übung verbessert das Lernen und die Hippocampus-Neurogenese bei älteren Mäusen. In: J. Neurosci. 25, 8680–8685. doi: 10.1523/JNEUROSCI.1731-05.2005
PubMed Zusammenfassung | CrossRef Volltext | Google Scholar
Verghese, J., Lipton, R. B., Katz, M. J., Hall, C. B., Dervy, C. A., Kuslansky, G., et al. (2003). Freizeitaktivitäten und das Demenzrisiko bei älteren Menschen. In: N. Engl. Dr. Med. 348, 2508–2516. doi: 10.1056/NEJMoa022252
PubMed Zusammenfassung | CrossRef Volltext / Google Scholar
Vital, T. M., Stein, A. M., de Melo Coelho, F. G., Arantes, F. J., Teodorov, E., Santos-Galduróz, R. F., et al. (2014). Körperliche Bewegung und vaskulärer endothelialer Wachstumsfaktor (VEGF) bei älteren Menschen: eine systematische Überprüfung. Bogen. Gerontologie. Geriatr. 59, 234–239. doi: 10.1016/j.archger.2014.04.011
PubMed Zusammenfassung / CrossRef Volltext / Google Scholar
Voelcker-Rehage, C., und Niemann, C. (2013). Strukturelle und funktionelle Veränderungen des Gehirns im Zusammenhang mit verschiedenen Arten von körperlicher Aktivität über die gesamte Lebensspanne. Neurowissenschaften. Biobehav. Offenbarung 37, 2268-2295. doi: 10.1016/j.neubiorev.2013.01.028
PubMed Abstract / CrossRef Full Text / Google Scholar
Zatorre, RJ, Felder, RD und Johansen-Berg, H. (2012). Plastizität in Grau und Weiß: Neuroimaging Veränderungen in der Gehirnstruktur während des Lernens. Nat. Neurowissenschaften. 15, 528–536. Ursprungsbezeichnung: 10.1038/nn.3045
PubMed Abstract | CrossRef Full Text | Google Scholar
Zimmermann, P., and Fimm, B. (2002). Testbatterie zur Aufmerksamkeitsprüfung (TAP), Version 1.7, Handbuch—Teil1. Würselen: Psytest.