
introduktion
den nuvarande demografiska förändringen i västerländska samhällen, som involverar både en relativ och en absolut ökning av antalet äldre människor, har väckt ökande vetenskapligt intresse för geriatriska problem. I detta sammanhang blir begreppen framgångsrik åldrande allt viktigare. Flera studier har visat att fysisk träning kan spela en nyckelroll i hälsosamt åldrande och i förebyggandet av kognitiv nedgång och neurodegenerativa sjukdomar (Colcombe et al., 2006; Erickson et al., 2011). Nya recensioner som sammanfattar epidemiologiska, tvärsnitts-och interventionsstudier stöder fysisk aktivitet som en gynnsam metod för att inducera neuroplasticitet i sen vuxen ålder (Gregory et al., 2012; Voelcker-Rehage och Niemann, 2013; Bamadis et al., 2014). Erickson et al. (2012) har dragit slutsatsen att högre kardiorespiratorisk kondition och aeroba fysiska aktivitetsnivåer är förknippade med större gråmaterialvolymer i prefrontala regioner och hippocampus. Men inte bara kardiovaskulär träning utan också koordinativ träning (Niemann et al., 2014) och kognitiv träning (Bamadis et al., 2014) har visat sig inducera gråmaterialplasticitet och förbättra kognitiva funktioner hos äldre vuxna.
djurforskning har föreslagit att en kombination av fysisk träning med sensorisk anrikning har den starkaste effekten på uppkomsten av nya neuroner—främst i hippocampus—och att endast denna kombination säkerställer den bestående överlevnaden hos de nyfödda cellerna (Kempermann et al., 1997; van Praag et al., 2005). Kattenstroth et al. (2013) har föreslagit att ”dansaktiviteter bör betraktas som en motsvarighet till berikade miljöförhållanden för människor eftersom de ger en individ ökade sensoriska, motoriska och kognitiva krav.”Trots detta uppmuntrande uttalande är studier som undersöker effekterna av dansträning på hjärnstruktur eller funktion knappa. Efter en 6-månaders dans intervention, Kattenstroth et al. (2010) har rapporterat betydande förbättringar i kognitiv, taktil och motorisk prestanda hos deltagande seniorer. Resultaten av en prospektiv studie av 469 personer äldre än 75 år under en median uppföljningsperiod på 5, 1 år har visat att dans är förknippad med en markant minskad risk för demens (Verghese et al., 2003). I alla fall, h Avsugningarfner et al. (2011) har rapporterat minskade volymer i flera hjärnregioner inklusive den främre hippocampala bildningen och delar av parieto-insulär vestibulär cortex hos professionella dansare och slackliners jämfört med icke-professionella. Detta resultat kan indikera att intensiv och repetitiv träning av samma motoriska färdigheter leder i samband med specialisering till minskade volymer i vissa hjärnregioner. I den här studien utformar vi därför ett speciellt dansträningsprogram som ständigt krävde att deltagarna lärde sig nya rörelsemönster (M., 2016). För att bedöma de specifika fördelarna med detta ingripande jämförde vi vårt nydesignade dansprogram med en aktiv snarare än en passiv kontrollgrupp, som deltog i ett konventionellt hälsosportprogram där deltagarna vanligtvis utförde repetitiva fysiska övningar som cykling på en ergometer. Dessutom, eftersom vi var intresserade av interventionernas tidsmässiga dynamik, bedömde vi effekterna på hjärnans struktur och funktion efter 6 och 18 månaders träning. På så sätt försökte vi bedöma om det är fördelaktigt att förlänga interventioner, eftersom vissa hjärnregioner kan kräva mer träning än andra, eller om det finns en gräns varefter mer träning blir skadlig. Slutligen, på jakt efter en potentiell mekanism som ligger bakom neuroplasticitet, mätte vi BDNF-nivåerna i perifert blod. Flera studier har föreslagit att BDNF främjar differentieringen av nya neuroner och synapser (Huang och Reichardt, 2001; Lessmann och Brigadski, 2009; Park och Poo, 2013; Edelmann et al., 2014). BDNF har därför föreslagits vara en medlare för neuroplasticitet hos vuxna (fl Jacobel et al., 2010).
material och metoder
deltagare och experimentell Design
studien utformades som en 18-månaders kontrollerad intervention. Studien godkändes av etikutskottet vid Otto-von-Guericke University, Magdeburg, och alla ämnen undertecknade en skriftlig informerad och de fick inte betalning för sitt deltagande. Sextiotvå friska äldre individer (63-80 år) rekryterade via meddelanden i lokala tidningar screenades för studien. Uteslutningskriterierna var klaustrofobi, tinnitus, metallimplantat, Tatueringar, diabetes mellitus, depression (Beck-Depressionsinventering, BDI-II > 13), kognitiva underskott (Mini-Mental tillståndsundersökning, MMSE < 27), neurologiska sjukdomar och regelbunden träning (1 h/vecka i genomsnitt). På grundval av dessa kriterier uteslöts 10 personer. De återstående 52 deltagarna tilldelades sedan slumpmässigt till antingen dansen eller sportgruppen genom att använda webbplatsen www.randomization.com och kontroll för ålder, MMSE status och fysisk kondition. Bedömningar utfördes vid baslinjen, efter 6 och efter 18 månaders träning (Figur 1). Tjugotvå deltagare slutförde hela interventionen och alla mätningar. Tabell 1 ger detaljerad demografisk data för dessa deltagare. Inga gruppskillnader avseende demografiska data hittades.
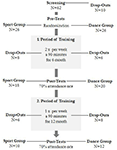
Figur 1. Flödesschema över deltagarnas rekrytering.
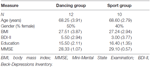
tabell 1. Demografisk information om deltagarna vid baslinjen.
interventioner
interventionerna separerades i två perioder. Under den första perioden tränade ämnena två gånger per vecka i 90-minuters sessioner i 6 månader. Av praktiska skäl (tillgång till deltagare och tränare) omfattade den andra 12-månadersperioden en minskad träningsfrekvens på en gång per vecka. Båda interventionsprogrammen utfördes i ett gruppsammanhang med musik för att kontrollera för psykosociala interaktioner. Villkorlig belastning undersöktes genom att registrera pulsvärdena under träningspasserna och genom att beräkna den individuella träningspulsen enligt Karvonen et al. (1957) med en faktor 0.6 för omfattande uthållighetsträning. Därför var de två träningsprogrammen jämförbara när det gäller intensitet, varaktighet och frekvens. Båda övervakades av erfarna instruktörer.
dansgrupp
deltagare i dansgruppen deltog i ett nydesignat träningsprogram där de ständigt ombads att lära sig nya rörelsesekvenser. Dessa koreografier krävde samordning av olika kroppsdelar (dvs ben, armar, bagageutrymme) i rymden under olika belastningsförhållanden (fysisk belastning, precision, situation och tidstryck). Ämnena var tvungna att lära sig koreografierna utantill, vilket också ställde höga krav på minnet. Programmet bestod av fem olika genrer (linedans, jazzdans, rock ”n” roll och square dance), som byttes efter var fjärde session. Under interventionens gång ökade koordinativa krav och tidspress genom att införa mer komplexa danssteg och koreografier och genom att öka slag per minut i musiken.
sportgrupp
deltagare i sportgruppen genomförde ett konventionellt styrketräningsprogram med huvudsakligen repetitiva övningar och låga krav på helkroppskoordination och minne. Varje session omfattade 20-min enheter av uthållighet, styrka-uthållighet och flexibilitetsträning. Uthållighetsträningen utfördes på cykel ergometrar. I styrka-uthållighetsenheten utfördes växlande rörelser (t.ex. bicepskrullar, knäböj, sit-ups), men komplexa helkroppsrörelser undviks för att hålla koordinativa krav låga. Flexibilitetsenheten bestod huvudsakligen av stretchövningar.
resultatmått
kardiovaskulär kondition
kardiovaskulär kondition bedömdes med det fysiska arbetsförmågan 130-testet (PWC 130). PWC 130 är uteffekten (mätt i watt) som en person kan uppnå på en cykelergometer under en hjärtfrekvens på 130 bpm. För beräkningar använde vi vilopuls och relativ fysisk kapacitet .
neuropsykologisk testning
ett omfattande batteri av neuropsykologiska tester utfördes på ämnena. För den aktuella studien rapporteras endast resultaten av ett verbalt kort – och långsiktigt minnestest, ”VLMT” (en anpassad tysk version av ”Rey auditive Verbal Learning Test”; Helmstaedter och Durwen, 1990) och ett uppmärksamhetstestbatteri (Test of Attentional Performance (TAP); Zimmermann och Fimm, 2002).
BDNF
fastande blodprover togs på morgonen av de neuropsykologiska bedömningarna. Från blodproverna bestämdes plasmakoncentrationerna av BDNF med sandwich ELISAs (BDNF-Duosetter; R&d-system, Wiesbaden, Tyskland) som tidigare beskrivits (Schega et al., 2016).
Mr
MR-bilder förvärvades på en 3 Tesla Siemens MAGNETOM Verio (Syngo MR B17) med användning av en 32-kanals huvudspole. Högupplösta T1-viktade mprage-sekvenser förvärvades med användning av ett 3D-magnetiseringsberedt snabbt gradientekobildningsprotokoll (224 sagittalskivor, voxelstorlek: 0,8 0,8 0,8 mm3, TR: 2500 ms, TE: 3,47 ms, TI: 1100 ms, flip-vinkel: 7 msk). MR-bilderna analyserades med hjälp av voxelbaserad morfometri (VBM) implementerad i SPM 12 (Welcome Department of Cognitive Neurology, London, Storbritannien). VBM är en helhjärnans opartisk teknik för analys av regional gråmaterialvolym och vävnadsförändringar (Ashburner och Friston, 2000).
förbehandling involverade gråmaterialsegmentering, mallskapande via DARTEL, rumslig normalisering till standardiserat Montreal Neurological Institute (MNI) utrymme och utjämning med en Gaussisk kärna av 8 mm Full bredd vid halv maximal (FWHM).
för att analysera skillnaden i gråmaterialvolymförändringar mellan grupper tillämpades en fullfaktoriell design med faktorgruppen (dans, sport) och tid (0, 6 och 18 månader). När det gäller betydande grupp-interaktioner mellan tidsinteraktioner i grupp, post hoc t-tester mellan på varandra följande par av tidpunkter (0 mot 6; 0 mot 18; 6 mot. 18 månader) beräknades separat för varje grupp. En tröskel för p < 0.001 (okorrigerad) tillämpades för alla analyser.
resultat
presentationen av resultaten är strukturerad enligt följande. Vi letade först efter en allmän interventionseffekt (faktortid) och letade sedan efter differentiella effekter av de två interventionerna (interaktionsgrupp bisexuell tid); slutligen utfördes en mer detaljerad analys av den tidsmässiga dynamiken via post hoc parvisa jämförelser.
kardiovaskulär kondition
kardiovaskulär kondition mätt med PWC 130 (Tabell 2, relativ fysisk kapacitet) skilde sig inte mellan grupperna vid baslinjen. Dessutom ökade PwC 130-poängen inte signifikant under hela tiden för någon av interventionerna.

tabell 2. Medel och (SD) för fitness, kognitiv funktion, BDNF-plasmanivåer och total gråmaterialvolym inom träningsgrupper över interventionen.
neuropsykologiska tester
en signifikant huvudeffekt av tiden observerades för verbalt korttidsminne (vlmt early recall; F(2,19) = 6.438, p = 0.004, oc-2 = 0.253), verbal långsiktig fri återkallelse (vlmt late recall; F(2,19) = 3.387, p = 0.049, oc-2 = 0.244), verbal långfristig återkallelse termigenkänning (vlmt-igenkänning; f(2,19) = 5.352, p = 0.009, oc 2 = 0.220) och UPPMÄRKSAMHETSREAKTIONSTID (Subtest flexibilitet; f(2,19) = 19.156, p < 0.001, oc 2 = 0.489). Inga signifikanta tidsinteraktioner för grupper av grupper uppstod.
post hoc parvisa jämförelser visade signifikanta förbättringar från baslinjen till 18 månader och från 6 till 18 månader i alla tre VLMT-underkategorierna i båda grupperna. När det gäller TAP-reaktionstider sågs en signifikant förbättring i jämförelsen av baslinjen till 18-månadersdata i båda grupperna.
BDNF
plasmanivåerna av BDNF analyserades i blodprover före träningens början samt 6 månader och 18 månader efter träningens början. Absoluta BDNF-plasmanivåer sammanfattas i Tabell 2. De intraindividuella förändringarna i BDNF-nivå avslöjade en signifikant ökning av dansgruppen, medan den intraindividuella BDNF-nivån förblev konstant efter 6 månaders träning i sportgruppen. Inga ytterligare förändringar i BDNF inträffade efter 18 månader i någon grupp (Figur 2).
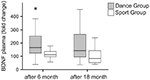
Figur 2. Intraindividuella förändringar i BDNF-plasmanivåer efter intervention. BDNF-plasmanivåer analyserades i blodprover av deltagare som utförde ett dansutbildningsprogram eller ett idrottsutbildningsprogram före träningens början, efter 6 månaders träning och efter 18 månaders träning. Den relativa ökningen av BDNF-nivåer kvantifierades. BDNF-nivåerna ökade signifikant i dansgruppen efter 6 månaders träning (Mann-Whitney U-test, p < 0.004) och sjönk nästan till baslinjen för förbehandlingsvärdet efter 18 månaders dansträning. Det var ingen förändring under hela tidskursen i sportgruppen (Friedman-testet, p = 0,319), medan en signifikant förändring under hela tidskursen observerades i dansgruppen (Friedman-testet, p = 0,028). Box tomter: minimum, 25: e percentilen, median, 75: e percentilen. * p 0,05 0,0.
MRI
en signifikant grupp av interaktion med tiden i den vänstra precentral gyrus och höger parahippocampus observerades. Post hoc t-test mellan baslinjen och 6-månadersdata visade en signifikant ökning av gråmaterialvolymen i vänster precentral gyrus av endast dansarna. I jämförelsen av baslinjen och 18-månadersdata utöver den precentrala gyrusen uppvisade dansarna en signifikant ökning av gråmaterialvolymen i rätt parahippocampal gyrus, vilket också var den enda signifikanta förändringen i intervallet från 6 till 18 månader. Således uppstod volymökningen i precentral gyrus efter 6 månader och förblev stabil under det återstående dansutbildningsintervallet, medan förändringen i parahippocampal gyrus inträffade endast under det senare träningsintervallet (Figur 3).
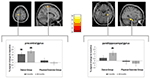
Figur 3. Tid efter gruppinteraktionsanalys, testning för större volymförändringar i dansen jämfört med sportgruppen. En signifikant ökning av grå materia hittades i precentral gyrus (Montreal Neurological Institute (MNI)-koordinater: x = -16; y = -18; z = 77) och i parahippocampal gyrus gyrus (MNI-koordinater: x = 34; y = -26; z = -20). Rutans tomter visar de relativa gråmaterialförändringarna i toppvoxeln. * p 0,05 0,0.
diskussion
i denna studie jämförde vi effekterna av deltagande i antingen ett dansprogram eller ett konventionellt sportprogram för fysisk kondition på hjärnfunktion och volym hos friska seniorer. Dansprogrammet var en nydesignad intervention som krävde ständigt att lära sig nya danskoreografier. Det konventionella sportprogrammet fokuserade främst på repetitiva motorövningar. Som ett huvudfynd observerade vi att efter 6 månaders träning hade volymerna i dansarnas vänstra precentrala gyrus ökat mer än i sportgruppen. Efter ytterligare 12 månaders träning observerades en ytterligare volymökning i dansarnas högra parahippocampala gyrus. BDNF-nivåerna ökade under de första 6 månaderna av dansträning och återvände till förbehandlingsvärdena efter 18 månader. I den konventionella sportgruppen var en liknande ökning av BDNF inte uppenbar. Eftersom de kardiovaskulära konditionsnivåerna under interventionerna förblev konstanta i båda grupperna, kunde de observerade effekterna inte hänföras till förbättringar i fysisk kondition utan verkade istället vara relaterade till dansprogrammets specifika egenskaper. Dessa funktioner inkluderade kravet att ständigt lära sig nya koreografier (dvs. minne), att integrera multisensorisk information, att samordna hela kroppen och att navigera i rymden.
hjärnförändringar
precentral gyrus är avgörande för kontrollen av frivilliga motorfunktioner. Ökningen av gråmaterialvolymen i den precentrala gyrusen i dansgruppen kan därför ha baserats på de komplexa och ständigt föränderliga rörelsemönster som dansarna var tvungna att utföra. Dessa rörelser krävde samtidig samordning av flera delar av kroppen i olika riktningar och anpassning till musikens varierande rytmer (polycentrisk och polyrytmisk). Reflektera över dessa komplexa samordningskrav, Brown et al. (2006) har rapporterat dansinducerade aktiveringar i putamen, den primära motorbarken och det kompletterande motorområdet (SMA), vilket visas av PET. Andra studier har indikerat en koppling mellan koordinationskrav (t.ex. balansering, jonglering) och neuroplasticitet i den precentrala regionen (Boyke et al., 2008; Taubert et al., 2010). Därför överensstämde den dansrelaterade volymökningen inom detta område med förväntningar baserade på litteraturen.
parahippocampal gyrus är en del av den yttre bågen i det limbiska systemet och spelar en viktig roll i arbetsminnet och episodisk minneshämtning (Pantel et al., 2003). Enligt Bliss och Lomo (1973) utgör parahippocampal gyrus gränssnittet mellan minnet och nutidens erfarenhetsmedvetande, eftersom det är sammankopplat av perforantkanalen både till regioner i frontalloben, som är associerade med arbetsminnet, och till hippocampus, den centrala strukturen i episodisk minneskodning och rumslig navigering. Många VBM-studier har rapporterat en åldersrelaterad volymförlust i parahippocampala regioner (Tisserand et al., 2002). Dessutom, Echuskvarri et al. (2011) har föreslagit att parahippocampal atrofi är en tidig biomarkör av AD. Observationen att engagera sig i ett dansprogram under en längre period kan inducera neuroplastiska processer i denna viktiga minnesregion, därför särskilt uppmuntrande när det gäller att utveckla förebyggande strategier.
Temporal dynamik av grå materia Hjärnplasticitet
den observerade volymen ökar i de två hjärnregionerna som utvecklats vid olika tidpunkter. Dans ledde till en volymökning i motorområdena efter 6 månader (Se även Rehfeld et al., 2016), som förblev stabil under de följande 12 månaderna. Den dansassocierade volymökningen i parahippocampal gyrus framkom senare och observerades endast i 18-månadersdata. Den olika tidsmässiga dynamiken i utvecklingen av de två hjärnregionerna kan vara relaterad till skillnader i de underliggande cellulära mekanismerna. Djurforskning har föreslagit att angiogenes och bildandet av nya dendriter (Thomas et al., 2012) förekommer snabbt, medan förändringar i neuropilen uppträder mycket långsammare (Black et al., 1990). Hos människor har snabba ökningar av gråmaterialvolymen i de prefrontala regionerna observerats efter bara 2 veckors motoriskt lärande hos yngre vuxna (Taubert et al., 2010). För att inducera neuroplasticitet i hippocampus har längre träningsperioder rekommenderats (Erickson et al., 2012; Niemann et al., 2014).
intressant är att de ovannämnda volymökningarna från vår studie står i kontrast till resultaten från H Askornggi et al. (2010), som avslöjade att professionella balettdansare har minskat gråmaterialvolymer i vänster premotorisk cortex, SMA, Putamen och superior frontal gyrus och resultaten av H Askorbfner et al. (2011), som rapporterar minskade volymer i flera hjärnregioner inklusive den främre hippocampalbildningen hos professionella dansare och slackliners jämfört med icke-professionella. Dessa studier var emellertid tvärsnittsobservationsstudier som jämförde hjärnorna hos professionella dansare med de hos icke-professionella. Dessutom bestod vår dansgrupp av gamla gamla nybörjare. Det har visats att inlärning av en ny färdighet först leder till rekrytering av ytterligare neurala resurser. Senare, när färdigheten blir mer automatisk, behövs mindre neuronala resurser vilket kan leda till volymminskningar hos dem med långvarig erfarenhet. Baserat på dessa initiala förhållanden (nybörjare, äldre vuxna) och vårt specialdesignade dansutbildningsprogram, som krävde ständigt nytt lärande av rörelsemönster, är det möjligt att effekterna av specialiseringsinducerad volym minskar som rapporterats av H Exceptionalnggi et al. (2010) observeras inte.
det anses allmänt att motorisk träning initialt inducerar hjärnvolymökningar. Långvarig träning leder emellertid till automatisering, vilket kan ha motsatta effekter på kortikalvolymen, eftersom mindre kortikal kontroll behövs efter att motoriken har blivit fullständigt etablerad (Taube, 2008). Vårt dansträningsprogram var därför speciellt utformat för att undvika sådan automatisering, vilket kan förklara varför, åtminstone inom 18 månader, inga kortikala volymminskningar observerades i vår studie.
kognitiva funktioner visade också olinjär utveckling, varigenom verbalt minne ökade endast under den andra träningsperioden. När det gäller uppmärksamhetsprestanda observerades signifikanta förbättringar efter bara 6 månader i båda grupperna (Rehfeld et al., 2016). Dessa resultat stöder tidigare rapporter om de positiva effekterna av fysiska ingrepp på neuropsykologiska tester (Bamadis et al., 2014). I kognitiv förmåga data, i motsats till hjärndata, uppstod emellertid inga gruppskillnader. Andra har rapporterat överlägsna effekter av kombinerad kognitiv och fysisk träning i motsats till enstaka ingrepp (Oswald et al., 2006). Vi kommer att utvidga våra insatser ytterligare för att testa om gruppskillnader i kognition kan uppstå vid ännu senare tidpunkter.
underliggande cellulära och molekylära mekanismer för Gråmaterialplasticitet
även om VBM är en bildmodalitet som avslöjar volymförändringar i hjärnan, tillåter denna teknik inte kausala slutsatser angående de underliggande neurofysiologiska processerna. Neurogenes, synaptogenes och angiogenes är bara några av de mekanismer som har föreslagits vara grunden för förändringar i hjärnvolymen (Zatorre et al., 2012). Som medlare av effekterna av kardiovaskulär kondition på hjärnan studeras tillväxtfaktorer som BDNF, insulinliknande tillväxtfaktor (IGF) och nervtillväxtfaktor (NGF) (Kirk-Sanchez och McGough, 2014). Men i vår studie, i motsats till tidigare (Erickson et al., 2011; Maass et al., 2015) fanns inga skillnader i kardiovaskulär kondition mellan grupper, och fitnessnivåerna förändrades inte under interventionerna. Den senare observationen var förmodligen relaterad till vår kontroll av den individuella hjärtfrekvensen, som vi syftade till att upprätthålla i den aeroba zonen. BDNF-förändringar har emellertid också associerats med fysisk aktivitet, social interaktion och positiv stress (Mattson, 2008), och inte alla studier har observerat en BDNF-ökning efter kardiovaskulär träning (Vital et al., 2014). Slutligen har djurforskning föreslagit att samordning men inte uthållighetsträning inducerar synaptogenes och glialförändringar (Black et al., 1990). Tillsammans kan de namngivna ytterligare faktorerna som driver BDNF-utsöndring ha varit mer avgörande under dans än under träningsaktiviteter, vilket förklarar varför endast dansare visade en BDNF-ökning under de första 6 månaderna. Observationen att BDNF-nivåerna återvände till baslinjen under de följande 12 månaderna medan volymökningar samtidigt observerades i parahippocampal gyrus indikerar dock att det måste finnas andra faktorer involverade i vuxen hjärnplasticitet än de som representeras av BDNF-nivåer i perifert blod.
När det gäller neurobiologiska mekanismer för utövad inducerad plasticitet också begreppet hjärnreserv (Satz et al., 2011) bör övervägas. Begreppet hjärnreserv beskriver individuella skillnader i en ökad baslinje adaptiv neuroplasticitet, vilket ger större dynamisk kapacitet för ombyggnad av kortikala kretsar till olika stressorer (Barulli och Stern, 2013; Freret et al., 2015).
perspektiv
resultaten av vår studie tyder på att en långsiktig dansingrepp kan vara överlägsen repetitiv fysisk träning för att inducera neuroplasticitet i den åldrande mänskliga hjärnan. Vi antar att denna fördel är relaterad till dansens multimodala natur, som kombinerar fysiska, kognitiva och koordinativa utmaningar. Så vitt vi vet är detta den första longitudinella, randomiserade studien som rekommenderar dansprogram som ett sätt att förhindra grå materia och kognitiv nedgång hos äldre. Ytterligare forskning behövs för att klargöra mer detaljerat den tidsmässiga dynamiken och de underliggande neurobiologiska mekanismerna för dansinducerad neuroplasticitet och om denna intervention verkligen har potential att minska risken för neurodegenerativa sjukdomar som Alzheimers.
Författarbidrag
PM utformade och utförde forskningen, analyserade data, skrev artikeln. KR designade och utförde forskningen. MS analyserade data. AH och VL utformade forskningen. MD: datainsamling, artikel revision. TB och JK utformade forskningen, analyserade data. NGM utformade forskningen, skrev artikeln.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
Ashburner, J. och Friston, K. J. (2000). Voxelbaserad morfometri-metoderna. Neuroimage 11, 805-821. doi: 10.1006 / nimg.2000.0582
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Bamadis, Pd, Vivas, AB, Styliadis, C., Frantzidis, M., Klados, M., Schlee, W., et al. (2014). En genomgång av fysiska och kognitiva ingrepp i åldrande. Neurovetenskap. Biobehav. Rev. 44, 206-220. doi: 10.1016/j.neubiorev.2014.03.019
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Barulli, D. Och Stern, Y. (2013). Effektivitet, kapacitet, kompensation, underhåll, plasticitet: nya koncept i cognitice reserve. Trender Cogn. Sci. 17, 502–509. doi: 10.1016/j.tics.2013.08.012
PubMed Abstrakt | CrossRef fulltext | Google Scholar
svart, Je, Isaacs, kr, Anderson, Bj, Alcantara, AA och Greenough, WT (1990). Lärande orsakar synaptogenes, medan motorisk aktivitet orsakar angiogenes, i cerebellär cortex hos vuxna råttor. Proc. Natl. Acad. Sci. USA 87, 5568-5572. doi: 10.1073/pnas.87.14.5568
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Bliss, TV och Lomo, T. (1973). Långvarig potentiering av synaptisk överföring i tandat hos den aneastetiserade kaninen efter stimulering av perforeringsvägen. J. Physiol. 232, 331–356. doi: 10.1113 / jphysiol.1973.sp010273
PubMed Abstract / CrossRef fulltext / Google Scholar
Boyke, J., Driemeyer, J., Gaser, C., B exceptional, C. och May, A. (2008). Träningsinducerad hjärnstruktur förändras hos äldre. J. Neurosci. 28, 7031–7035. doi: 10.1523/JNEUROSCI.0742-08.2008
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Brown, S., Martinez, M. och Parsons, M. (2006). Den neuronala grunden för mänsklig dans. Cereb. Cortex 16, 1157-1167. doi: 10.1093 / cercor / bhj057
PubMed Abstrakt / CrossRef fulltext/Google Scholar
Colcombe, sj, Erickson, ki, Scalf, Pe, Kim, JS, Prakash, R., McAuley, E., et al. (2006). Aerob träning ökar hjärnvolymen hos åldrande människor. J. Gerontol. A. Biol. Sci. Med. Sci. 61, 1166–1170. doi: 10.1093 / gerona / 61.11.1166
PubMed Abstract | CrossRef fulltext | Google Scholar
Echcorivarri, C., Aalten, P., Uyling, HBM, Jacobs, H. I. L., Visser, PJ, Gronenschild, M., et al. (2011). Atrofi i parahippocampal gyrus som en tidig biomarkör av Alzheimers sjukdom. Hjärnstruktur. Funct. 215, 265–271. doi: 10.1007 / s00429-010-0283-8
PubMed Abstract / CrossRef fulltext / Google Scholar
Edelmann, E., Lessmann, V. och Brigadski, T. (2014). Pre – och postsynaptiska vridningar i BDNF-utsöndring och verkan i synaptisk plasticitet. Neurofarmakologi 76, 610-627. doi: 10.1016 / j. neuropharm.2013.05.043
PubMed Abstrakt / CrossRef fulltext/Google Scholar
Erickson, K. I., Voss, MW, Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., et al. (2011). Träning ökar storleken på hippocampus och förbättrar minnet. Proc. Natl. Acad. Sci. USA 108, 3017-3022. doi: 10.1073/pnas.1015950108
PubMed Abstrakt / CrossRef fulltext/Google Scholar
Erickson, ki, Weinstein, AM och Lopez, ol (2012). Fysisk aktivitet, hjärnplasticitet och Alzheimers sjukdom. Båge. Med. Res 43, 615-621. doi: 10.1016/j.arcmed.2012.09.008
PubMed Abstract / CrossRef fulltext / Google Scholar
fl Jacobel, A., Ruscheweyh, R., kr., K., Willemer, C., Vinter, B., V. (2010). Fysisk aktivitet och minnesfunktioner: är neurotrofiner och cerebral gråmaterialvolym den saknade länken? Neuroimage 49, 2756-2763. doi: 10.1016/j.neuroimage.2009.10.043
PubMed Abstract | CrossRef fulltext | Google Scholar
Freret, T., Gaudreau, P., Schumann-Bard, P., Billard, JM och Popa-Wagner, A. (2015). Mekanismer som ligger till grund för den neuroprotektiva effekten av hjärnreserv mot depression i slutet av livet. J. Neural. Transm. (Wien) 122, S55–S61. doi: 10.1007 / s00702-013-1154-2
PubMed Abstract / CrossRef fulltext/Google Scholar
Gregory, SM, Parker, B. och Thompson, Pd (2012). Fysisk aktivitet, kognitiv funktion och hjärnhälsa: vad är träningsutbildningens roll för att förebygga demens? Hjärnan Sci. 2, 684–708. doi: 10.3390 / brainsci2040684
PubMed Abstrakt / CrossRef fulltext / Google Scholar
h Jacobnggi, J., Koeneke, S., Bezzola, L., Och J Jacobncke, L. (2010). Strukturell neuroplasticitet i det sensorimotoriska nätverket av professionella kvinnliga balettdansare. Brum. Hjärna Mapp. 31, 1196–1206. doi: 10.1002 / hbm.20928
PubMed Abstrakt / CrossRef fulltext/Google Scholar
Helmstaedter, C. och Durwen, HF (1990). Det verbala inlärnings-och retentionstestet. Ett användbart och differentierat verktyg för att utvärdera verbalt minnesprestanda. Schweiz. Båge. Neurol. Psykiatriker. (1985) 141, 21–30.
PubMed Abstract / Google Scholar
Huang, Ej och Reichardt, LF (2001). Neurotrofiner: roller i neuronal utveckling och funktion. Annu. Rev Neurosci. 24, 677–736. doi: 10.1146/annurev.neuro.24.1.677
PubMed Abstract | CrossRef fulltext | Google Scholar
H C., Binetti, C., Hamilton, D. A., Stephan, T., Flanagin, V. L., Linn, J., et al. (2011). Strukturell och funktionell plasticitet av hippocampalformationen i professionella dansare och slackliners. Hippocampus 21, 855-865. doi: 10.1002/hipo.20801
PubMed Abstract / CrossRef fulltext / Google Scholar
Karvonen, M. J., Kentala, E., Och Mustala, O. (1957). Effekterna av träning på hjärtfrekvens: en longitudinell studie. Ann. Med. Exp. Biol. Fenn. 35, 307–315.
PubMed Abstract / Google Scholar
Kattenstroth, JC, Kalisch, T., Holt, S., Tegethoff, M. och Dinse, HR (2013). Sex månaders dansintervention förbättrar postural, sensorimotorisk och kognitiv prestanda hos äldre utan att påverka hjärt-respiratoriska funktioner. Front. Åldrande Neurosci. 5:5. doi: 10.3389/fnagi.2013.00005
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Kattenstroth, J. C., Kolankowska, I., Kalisch, T. och Dinse, HR (2010). Överlägsen sensorisk, motorisk och kognitiv prestanda hos äldre individer med fleråriga dansaktiviteter. Front. Åldrande Neurosci. 2:31. doi: 10.3389/fnagi.2010.00031
PubMed Abstrakt / CrossRef fulltext/Google Scholar
Kempermann, G., Kuhn, Hg och Gage, fh (1997). Fler hippocampala neuroner hos vuxna möss som lever i en berikad miljö. Natur. 386, 493–495. doi: 10.1038 / 386493a0
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Kirk-Sanchez, NJ, och McGough, El (2014). Fysisk träning och kognitiv prestanda hos äldre: nuvarande perspektiv. Clin. Interv. Åldrande 9, 51-62. doi: 10.2147 / CIA.S39506
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Lessmann, V. och Brigadski, T. (2009). Mekanismer, platser och kinetik för synaptisk BDNF-sekretion: en uppdatering. Neurovetenskap. Res 65, 11-22. doi:10.1016 / j. neures.2009.06.004
PubMed Abstract / CrossRef fulltext/Google Scholar
Maass, A., D. A., S., Goerke, M., Becke, A., Sobieray, U., Neumann, K., et al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M. A., Kaufmann, J., et al. (2016). Effekter av dans-och hälsosportträning på den grå hjärnämnet hos friska seniorer. Sportwiss. 46, 213-222. doi: 10.1007 / s12662-016-0411-6
CrossRef fulltext / Google Scholar
Niemann, C., Godde, B. Och Voelcker-Rehage, C. (2014). Inte bara kardiovaskulär, men också samordnad träning ökar hippocampalvolymen hos äldre vuxna. Front. Åldrande Neurosci. 6:170. doi: 10.3389/fnagi.2014.00170
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Oswald, W. D., Gunzelmann, T., Rupprecht, R. och Hagen, B. (2006). Differentiella effekter av singel kontra kombinerad kognitiv och fysisk träning med äldre vuxna: SimA-studien i ett 5-årigt perspektiv. EUR. J. Åldrande 3, 179-192. doi: 10.1007 / s10433-006-0035-z
CrossRef fulltext / Google Scholar
Pantel, J., Kratz, B., Essig, M., och Schr Xhamster, J. (2003). Parahippocampal volymunderskott hos personer med åldrande associerad kognitiv nedgång. Är. J. Psykiatri 160, 379-382. doi: 10.1176/appi.ajp.160.2.379
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Park, H., och Poo, M. M. (2013). Neurotrofinreglering av nervkretsutveckling och funktion. Nat. Rev Neurosci. 14, 7–23. doi: 10.1038 / nrn3379
PubMed Abstract / CrossRef fulltext / Google Scholar
Rehfeld, K., H Jacobkelmann, A., Kaufmann, J., och M Jacobller, ng (2016). ”Effekter av en dans vs. en fitnessträning på hjärnplasticitet, balansprestanda och uppmärksamhet hos friska seniorer: ett nytt tillvägagångssätt med SPM 12 för parvis longitudinell gruppjämförelse,” in Proceeding-International Conference of Sports and Neuroscience (Magdeburg), 127-138.
Satz, P., Cole, M. A., Hardy, D. J. och Rassovsky, Y. (2011). Hjärna och kognitiv reserv: medlare (er) och konstruera validitet, en kritik. J. Clin. Exp. Neuropsychol. 33, 121–130. doi: 10.1080 / 13803395.2010.493151
PubMed Abstract | CrossRef fulltext/Google Scholar
Schega, L., Peter, B., Brigadski, T., Le Jacobmann, V., Isermann, B., Hamacher, D., et al. (2016). Effekt av intermittent normobarisk hypoxi på aerob kapacitet och kognitiv funktion hos äldre. J. Sci. Med. Sport 19, 941-945. doi: 10.1016/j.jsams.2016.02.012
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Taube, W. (2008). Neurofysiologiska anpassningar som svar på balansträning. Dtsch. Z. Sportmed. 63, 273–277. doi: 10.5960 / dzsm.2012.030
CrossRef fulltext / Google Scholar
Taubert, M., Draganski, B., Anwander, A., M Obbller, K., Horstmann, A., Villringer, A., et al. (2010). Dynamiska egenskaper hos mänsklig hjärnstruktur: inlärningsrelaterade förändringar i kortikala områden och tillhörande fiberanslutningar. J. Neurosci. 30, 11670–11677. doi: 10.1523/JNEUROSCI.2567-10.2010
PubMed Abstract / CrossRef fulltext / Google Scholar
Thomas, A. G., Dennis, A., Bandettini, P. A. och Johansen-Berg, H. (2012). Effekterna av aerob aktivitet på hjärnans struktur. Front. Psychol. 3:86. doi: 10.3389/fpsy.2012.00086
PubMed Abstrakt | CrossRef fulltext | Google Scholar
Tisserand, D. J., Pruessner, J. C., Sanz Arigita, Ej, van Boxtel, M. P., Evans, A. C., Jolles, J., et al. (2002). Regionala frontala kortikala volymer minskar differentiellt i åldrandet: en MR-studie för att jämföra volymetriska tillvägagångssätt och voxelbaserad morfometri. Neuroimage 17, 657-669. doi: 10.1016 / s1053-8119 (02) 91173-0
PubMed Abstrakt | CrossRef fulltext | Google Scholar
van Praag, H., Shubert, T., Zhao, C. och Gage, fh (2005). Övning förbättrar inlärning och hippocampal neurogenes hos åldrade möss. J. Neurosci. 25, 8680–8685. doi: 10.1523/JNEUROSCI.1731-05. 2005
PubMed Abstract / CrossRef fulltext/Google Scholar
Verghese, J., Lipton, RB, Katz, MJ, Hall, CB, Dervy, ca, Kuslansky, G., et al. (2003). Fritidsaktiviteter och risken för demens hos äldre. N. Engl. J. Med. 348, 2508–2516. doi: 10.1056/NEJMoa022252
PubMed Abstract | CrossRef fulltext/Google Scholar
Vital, T. M., Stein, A. M., De Melo Coelho, F. G., Arantes, F. J., Teodorov, E., Santos-Galdur Jacobz, R. F., et al. (2014). Fysisk träning och vaskulär endoteltillväxtfaktor (VEGF) hos äldre: en systematisk granskning. Båge. Gerontol. Geriatr. 59, 234–239. doi: 10.1016/j.archger.2014.04.011
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Voelcker-Rehage, C., och Niemann, C. (2013). Strukturella och funktionella hjärnförändringar relaterade till olika typer av fysisk aktivitet över hela livslängden. Neurovetenskap. Biobehav. Rev. 37, 2268-2295. doi: 10.1016/j.neubiorev.2013.01.028
PubMed Abstract / CrossRef fulltext / Google Scholar
Zatorre, Rj, Fields, rd och Johansen-Berg, H. (2012). Plasticitet i grått och vitt: neuroimaging förändringar i hjärnans struktur under inlärning. Nat. Neurovetenskap. 15, 528–536. doi: 10.1038 / nn.3045
PubMed Abstract / CrossRef fulltext / Google Scholar
Zimmermann, P. och Fimm, B. (2002). Testbatteri för uppmärksamhetstestning( TAP), version 1.7, Manuell—Del1. W Ubicrselen: Psytest.