
Introduction
高齢者の数の相対的および絶対的な増加を含む西洋社会の現在の人口統計学的変化は、高齢者の問題に対する科学的関心の高まりを引き起こしている。 この文脈では、成功した老化の概念はますます重要になってきています。 いくつかの研究は、身体運動が健康な老化および認知低下および神経変性疾患の予防において重要な役割を果たし得ることを示している(Colcombe et al. ら,2 0 0 6;Erickson e t a l., 2011). 疫学的、横断的および介入的研究を要約した最近のレビューは、成人後期に神経可塑性を誘導するための好都合な方法としての身体活動を支持する(Gregory et al. ら、2 0 1 2;Voelcker−RehageおよびNiemann、2 0 1 3;Bamadis e t a l., 2014). エリクソン他 (2012)は、より高い心呼吸フィットネスと好気性の身体活動レベルは、前頭前野および海馬におけるより大きな灰白質体積と関連していると結論付けてい しかし、心血管フィットネスだけでなく、調整運動(Niemann et al. ら、2 0 1 4)および認知訓練(Bamadis e t a l.、2014)は、高齢者における灰白質の可塑性を誘導し、認知機能を強化することが示されている。
動物の研究は、身体運動と感覚濃縮の組み合わせが、主に海馬における新しいニューロンの発生に最も強い影響を及ぼし、この組み合わせだけが新生児細胞の永続的な生存を保証することを示唆している(Kempermann et al. ら、1 9 9 7;van Praag e t a l., 2005). Kattenstroth et al. (2013)は、”ダンス活動は、感覚、運動、認知の要求が増加する個人を提供するため、人間の豊かな環境条件と同等とみなされるべきである”と示唆している。「この奨励的な声明にもかかわらず、ダンストレーニングが脳の構造や機能に及ぼす影響を調べる研究はほとんどありません。 6ヶ月のダンス介入の後、Kattenstroth et al. (2010)参加高齢者における認知、触覚および運動性能の有意な改善を報告している。 75歳以上の469人の被験者の前向き研究の結果は、5.1年のフォローアップ期間の中央値にわたって、ダンスが認知症の著しく減少したリスクと関連していることを示している(Verghese et al., 2003). しかし、Hüfner et al. (2011)は非専門家と比較される専門のダンサーおよびslacklinersの前方の海馬の形成そしてparieto島の前庭皮質の部分を含む複数の頭脳の地域の減らされた容積を報告し この知見は、同じ運動能力の集中的かつ反復的な訓練が、専門化の文脈でいくつかの脳領域の量の減少につながることを示している可能性がある。 したがって、本研究では、参加者が常に新しい動きパターンを学ぶことを必要とする特別なダンストレーニングプログラムを設計します(Müller et al., 2016). この介入の具体的な利点を評価するために、我々は、参加者は、通常、エルゴメーター上で自転車などの反復的な物理的な練習を行った従来の健康スポーツフィットネスプログラムに参加したアクティブではなく、受動的な対照群に私たちの新たに設計されたダンスプログラムを比較した。 さらに、介入の時間的ダイナミクスに興味があったため、6ヶ月と18ヶ月の訓練の後に脳の構造と機能に及ぼす影響を評価しました。 そうすることで、我々は、いくつかの脳領域が他の領域よりも多くの訓練を必要とする可能性があるため、介入を拡張することが有益であるかどうか、ま 最後に、神経可塑性の根底にある潜在的なメカニズムを求めて、我々は末梢血中のBDNFレベルを測定した。 いくつかの研究は、BDNFが新しいニューロンおよびシナプスの分化を促進することを示唆している(Huang and Reichardt、2001;Lessmann and Brigadski、2009;Park and Poo、2013;Edelmann et al., 2014). したがって、BDNFは、成体神経可塑性のメディエーターであることが提案されている(Floel e t a l., 2010).
材料と方法
参加者と実験デザイン
この研究は、18ヶ月の制御された介入として設計されました。 この研究は、マクデブルクのオットー-フォン-ゲリッケ大学の倫理委員会によって承認され、すべての被験者は書面に署名し、参加の支払いを受けなかった。 地元の新聞での発表を介して募集六十から二健康な高齢者(63-80歳)は、研究のためにスクリーニングされました。 除外基準は、閉所恐怖症、耳鳴り、金属インプラント、入れ墨、糖尿病、うつ病(Beck-Depressions Inventory、BDI-II>13)、認知障害(ミニ精神状態検査、MMSE<27)、神経学的疾患および定期的な運動(≥1時間/週)であった。 これらの基準に基づいて、10人の被験者を除外した。 残りの52人の参加者は、ウェブサイトを使用して、ダンスまたはスポーツグループのいずれかに無作為に割り当てられましたwww.randomization.com そして年齢、MMSEの状態および体力のための制御。 評価は、6ヶ月後および18ヶ月後のトレーニングのベースラインで実施された(図1)。 二十から二の参加者は、全体の介入とすべての測定を完了しました。 表1は、これらの参加者の詳細な人口統計データを示しています。 人口統計データに関するグループの違いは見つかりませんでした。
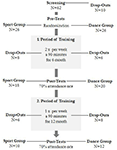
図1. 参加者募集のフローチャート。テーブル1
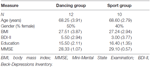
ベースラインでの参加者の人口統計情報。
介入
介入は二つの期間に分けられた。 最初の期間では、被験者は90分のセッションで週に二回6ヶ月間訓練を受けました。 実際的な理由(参加者とトレーナーの可用性)のために、第二の12ヶ月の介入期間は、週に一度のトレーニング頻度の減少を構成しました。 両方の介入プログラムは、心理社会的相互作用を制御するために、音楽とのグループコンテキストで実行されました。 条件付き負荷は、訓練セッション中のパルス値を記録することによって、およびKarvonen et alに従って個々の訓練心拍数を計算することによって調べた。 1957年(昭和20年)には、大規模な持久力トレーニングのための係数0.6を持つ。 したがって、2つの訓練プログラムは、強度、期間および頻度の点で同等であった。 両方とも経験豊富なインストラクターによって監督されました。
ダンスグループ
ダンスグループの参加者は、彼らが常に新しい動きのシーケンスを学ぶように求められた新たに設計されたトレーニングプロ これらの振り付けは、異なるひずみ条件(物理的なひずみ、精度、状況、時間の圧力)の下で、空間内の異なる身体部分(すなわち、脚、腕、胴)の調整を必要とした。 被験者は心で振り付けを学ばなければならなかったので、記憶にも高い要求を課しました。 プログラムは、5つの異なるジャンル(ラインダンス、ジャズダンス、ロックンロール、スクエアダンス)で構成され、4回目のセッションごとに切り替えられました。 介入の過程で、より複雑なダンスの動きや振り付けを導入し、音楽の分あたりのビートを増やすことによって、調整の要求と時間の圧力が増加しました。
スポーツグループ
スポーツグループの参加者は、主に反復的な練習と全身の調整と記憶の面で低い要求を持つ従来の強持久力トレーニングプロ 各セッションは、持久力、強さ-持久力と柔軟性のトレーニングの20分単位で構成されていました。 サイクルエルゴメーターで持久力トレーニングを行った。 強度耐久ユニットでは、交互の動き(例えば、上腕二頭筋カール、スクワット、腹筋)が行われたが、複雑な全身の動きは、調整要求を低く保つために避けられた。 柔軟性の単位は主に伸張の練習から成っていた。
アウトカム測定
心血管フィットネス
心血管フィットネスは、物理的な作業能力130テスト(PWC130)によって評価されました。 PWC130は、被験者が130bpmの心拍数の下でサイクルエルゴメーターで達成することができる電力出力(ワット単位で測定)です。 計算には、安静時心拍数と相対的な身体能力を使用しました。
神経心理学的検査
神経心理学的検査の広範なバッテリーは、被験者に行われました。 現在の研究の目的のために、口頭短期および長期記憶テスト、”VLMT”(”Rey聴覚口頭学習テスト”の適応ドイツ語版;HelmstaedterおよびDurwen、1990)および注意テストバッテリー(Attentional Performance(TAP)のテスト;ZimmermannおよびFimm、2002)の結果のみが報告されている。
BDNF
神経心理学的評価の朝に空腹時の血液サンプルを採取した。 血液サンプルから、BDNFの血漿濃度を、以前に記載されたように、サンドイッチElisa(BDNF Duosets;R<div i d=”1 1 7 8 8 6 3a9c”></div>D Systems,Wiesbaden,Germany)によって決定した(Schega e t a l., 2016).
MRI
MR画像は、3テスラSiemens MAGNETOM Verio(Syngo MR B17)で32チャンネルのヘッドコイルを使用して取得しました。 高分解能T1重み付けMPRAGE配列は、3D磁化調製されたrapid gradient echo imaging protocol(2 2 4個の矢状スライス、ボクセルサイズ:0. MR画像は、SPM1 2(Welcome Department o f Cognitive Neurology,London,UK)に実装されたボクセルベースの形態測定(VBM)を使用して分析した。 VBMは、局所的な灰白質体積および組織変化の分析のための全脳公平な技術である(AshburnerおよびFriston、2000)。
前処理には、灰白質セグメンテーション、DARTELを介したテンプレート作成、標準化されたモントリオール神経研究所(MNI)空間への空間正規化、最大半分(FWHM)で全幅8mmのガウスカーネルによる平滑化が含まれていた。
グループ間の灰白質体積変化の差を分析するために、因子グループ(ダンス、スポーツ)および時間(0、6および18ヶ月)を用いた完全階乗計画を適用した。 有意なグループ×時間相互作用の場合、時間点の連続したペア間の事後t検定(0対6;0対18;6対18;6対18;6対18;6対18;6対18;6対18;6対18;6対18;6対 18ヶ月)は、各グループについて別々に計算した。 Pのしきい値<0.001(訂正されていない)は、すべての分析に適用されました。
結果
結果の表示は次のように構成されています。 最初に一般的な介入効果(因子時間)を探し、次に二つの介入の差動効果(相互作用グループ×時間)を探し、最後に、時間的ダイナミクスのより詳細な分析は、ポスPWC130(表2、相対的な身体能力)によって測定された心血管フィットネスは、ベースラインでのグループ間で異ならなかった。
心血管フィットネス
さらに、PWC130スコアは、いずれの介入の時間経過を通じて有意に増加しなかった。表2

介入に関するトレーニンググループ内のフィットネス、認知機能、BDNF血漿レベルおよび総灰白質体積のための手段および(SD)。時間の有意な主な効果は、口頭短期記憶(VLMT早期リコール;F(2,19)=6.438、p=0.004、σ2=0.253)、口頭長期自由リコール(VLMT後期リコール;f(2,19)=3.387、p=0.049、σ2=0.244)、口頭長期認識19.156,p<0.001,λ2=0.489)。 重要な時間×グループの相互作用は現れませんでした。
ポストホックペアワイズ比較は、両方のグループのすべての三つのVLMTサブカテゴリでベースラインから18ヶ月と6から18ヶ月に有意な改善を示した。 タップ反応時間に関しては、両方のグループの18ヶ月のデータとベースラインの比較で有意な改善が見られた。
BDNF
BDNFの血漿レベルは、訓練開始前および訓練開始後6ヶ月および18ヶ月の血液サンプルで分析した。 絶対BDNF血漿レベルを表2に要約する。 BDNFレベルの個人内の変化は、ダンスグループの有意な増加を明らかにしたが、個人内BDNFレベルは、スポーツグループの訓練の6ヶ月後に一定のままであった。 BDNFのさらなる変化は、いずれのグループでも18ヶ月後には起こらなかった(図2)。
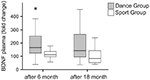
介入後のBDNF血漿レベルの個々の変化。 BDNF血漿レベルは、訓練の開始前、訓練の6ヶ月後および訓練の18ヶ月後に、ダンス訓練プログラムまたはスポーツ訓練プログラムを実施する参加者の血 BDNFレベルの相対的増加を定量化した。 BDNFレベルは、6ヶ月のトレーニング(Mann-Whitney U-test、p<0.004)後にダンスグループで有意に増加し、ダンストレーニングの18ヶ月後に前処理値のベースライン スポーツグループでは全時間コースにわたって変化はなかった(フリードマン検定、p=0.319)が、ダンスグループでは全時間コースにわたって有意な変化が観察された(フリードマン検定、p=0.028)。 箱ひげ図:最小、25パーセンタイル、中央値、75パーセンタイル。 *p≤0.05。
MRI
有意なグループ×時間相互作用は、左中心前回と右の海馬傍で観察されました。 ベースラインと6ヶ月のデータの間のポストhoc tテストは、ダンサーのみの左前中心回における灰白質体積の有意な増加を示した。 ベースラインと18ヶ月のデータの比較では、前中心回に加えて、ダンサーはまた、6から18ヶ月の間隔で唯一の有意な変化であった右傍海馬回の有意な灰白質の容積の増加を示した。 したがって、前中心回の体積増加は6ヶ月後に現れ、残りのダンストレーニング間隔にわたって安定したままであったが、後のトレーニング間隔でのみ傍海馬回の変化が起こった(図3)。

図3. グループの相互作用分析による時間、スポーツグループと比較してダンスの大きな音量の変化をテストします。 灰白質の有意な増加は、前中心回(モントリオール神経研究所(MNI)-座標:x=-16;y=-18;z=77)と傍海馬回(MNI-座標:x=34;y=-26;z=-20)で発見された。 箱ひげ図は、ピークボクセル内の相対的な灰白質の変化を示しています。 *p≤0.05。
ディスカッション
本研究では、ダンスプログラムまたは従来の体力スポーツプログラムのいずれかへの参加が健康な高齢者の脳機能 ダンスプログラムは、常に新しいダンスの振り付けを学ぶ必要が新たに設計された介入でした。 従来のスポーツプログラムは、主に反復運動に焦点を当てました。 主な発見として、我々は訓練の6ヶ月後に、ダンサーの左中心前回のボリュームがスポーツグループのものよりも増加していたことを観察した。 トレーニングの別の12ヶ月後、追加のボリュームの増加は、ダンサーの右傍海馬回で観察されました。 BDNFのレベルはダンスの訓練の最初の6か月の間に増加し、18か月後に前処理の価値に戻った。 従来のスポーツグループでは、BDNFの同様の増加は明らかではなかった。 介入の過程における心血管フィットネスレベルは両群で一定であったため,観察された効果は体力の改善に起因するものではなく,ダンスプログラムの特定の特徴に関連していると考えられた。 これらの機能には、常に新しい振り付け(すなわち、メモリ)を学び、多感覚情報を統合し、全身を調整し、空間内をナビゲートする必要が含まれていました。
脳の変化
前中心回は、自発的な運動機能の制御に不可欠です。 したがって,ダンスグループの前中心回における灰白質体積の増加は,ダンサーが実行しなければならなかった複雑で刻々と変化する動きパターンに基づいている可能性がある。 これらの動きは、身体のいくつかの部分を異なる方向に同時に調整し、音楽の様々なリズム(多中心性および多リズム性)に調整する必要があった。 これらの複雑な調整要件を反映して、Brown et al. (2006)は、petによって示されるように、被殻、一次運動野および補助運動領域(SMA)におけるダンス誘発活性化を報告している。 他の研究では、前中心領域における協調要求(例えば、バランス、ジャグリング)と神経可塑性との間の関連性が示されている(Boyke et al. ら、2 0 0 8;Taubert e t a l., 2010). したがって、この分野におけるダンス関連のボリュームの増加は、文献に基づく期待と一致していた。
傍海馬回は、辺縁系の外側弧の一部であり、作業記憶およびエピソード記憶検索において重要な役割を果たす(Pantel et al., 2003). Bliss and Lomo(1973)によると、傍海馬回は、作業記憶に関連する前頭葉の領域と、エピソード記憶のエンコードと空間ナビゲーションの中心構造である海馬の両方に穿孔路によって相互接続されているため、記憶と現在の経験的意識との間のインターフェイスを構成しています。 多くのVBM研究は、海馬傍領域における加齢に関連した体積損失を報告している(Tisserand e t a l., 2002). さらに、Echávarri et al. (2011)は、海馬傍萎縮がADの初期のバイオマーカーであることを示唆している。 長い期間のためのダンスプログラムに従事することは、この重要な記憶領域における神経形成プロセスを誘導することができるという観察,したがって、特に予防戦略の開発の面で奨励.
灰白質脳可塑性の時間的ダイナミクス
異なる時間に開発された二つの脳領域で観察されたボリュームの増加。 ダンスは6ヶ月後に運動領域の容積の増加をもたらした(Rehfeld et al.(2016年12月現在)。 傍海馬回のダンス関連ボリュームの増加は、後で浮上し、唯一の18ヶ月のデータで観察されました。 二つの脳領域の進化における異なる時間的ダイナミクスは、基礎となる細胞機構の違いに関連している可能性がある。 動物研究は、血管新生および新しい樹状突起の形成が示唆されている(Thomas e t a l. 2012年)は急速に起こるが、ニューロピルの変化ははるかに遅く起こる(Black et al., 1990). ヒトでは、若年成人における運動学習のわずか2週間後に、前頭前野領域における灰白質体積の急速な増加が観察されている(Taubert e t a l., 2010). 海馬において神経可塑性を誘導するために、より長い訓練期間が推奨されている(Erickson e t a l. ら、2 0 1 2;Niemann e t a l., 2014).興味深いことに、我々の研究からの前述の体積の増加は、Hänggi et al. (2010)は、プロのバレエダンサーが左運動前皮質、SMA、被殻および上前頭回における灰白質量を減少させていることを明らかにし、Hüfner et al. (2011)は、非専門家と比較してプロのダンサーやスラックライナーにおける前海馬形成を含むいくつかの脳領域におけるボリュームの減少を報告しています。 しかし、これらの研究は、プロのダンサーの脳を非専門家の脳と比較した横断的な観察研究であった。 また、私たちのダンスグループは、古い高齢者の初心者で構成されていました。 新しいスキルの学習は、最初に追加の神経リソースの募集につながることが示されています。 その後、スキルがより自動化されると、より少ない神経リソースが必要とされ、長期的な経験を持つ人の量が減少する可能性があります。 これらの初期条件(初心者、高齢者)と運動パターンの常に新しい学習を必要とする私たちの特別な設計されたダンストレーニングプログラムに基づいて、Hänggiら (2010)は観測されていない。
一般的に、運動訓練は最初に脳の容積の増加を誘導すると考えられている。 しかし、運動能力が完全に確立された後は皮質制御が必要ではないため、長時間の訓練は自動化につながり、皮質容積に逆の影響を及ぼす可能性があ したがって、私たちのダンストレーニングプログラムは、このような自動化を避けるために特別に設計されており、少なくとも18ヶ月以内に皮質体積の減少が私たちの研究で観察されなかった理由を説明するかもしれません。
認知機能も非線形発達を示し、それによって口頭記憶は第二の訓練期間中にのみ増加した。 注意性能に関しては、両方の群でわずか6ヶ月後に有意な改善が観察された(Rehfeld et al., 2016). これらの知見は、神経心理学的検査に対する物理的介入の有益な効果に関する以前の報告を支持する(Bamadis et al., 2014). しかし、認知能力データでは、脳データとは対照的に、グループの違いは現れなかった。 他の人たちは、単一の介入とは対照的に、認知的および身体的訓練の組み合わせの優れた効果を報告している(Oswald et al., 2006). 私たちは、認知のグループの違いがさらに後の時点で現れるかどうかをテストするために、介入をさらに拡張します。
灰白質可塑性の基礎となる細胞および分子メカニズム
VBMは、脳の体積変化を明らかにするイメージングモダリティですが、この技術は、基になる神経生理学的プロセスに関する因果的な結論を可能にしません。 神経発生、シナプス形成および血管形成は、脳容積変化の基礎であることが示唆されている機構のほんの一部である(Zatorre e t a l., 2012). 脳に対する心血管適応度の影響のメディエーターとして、BDNF、インスリン様成長因子(IGF)および神経成長因子(NGF)などの成長因子が研究されている(Kirk-Sanchez and McGough、2014)。 しかしながら、我々の研究では、以前のものとは対照的に(Erickson e t a l. ら、2 0 1 1;Maass e t a l.、2015)、心血管の適応度に差は群間に存在せず、適応度レベルは介入中に変化しなかった。 後者の観察は、おそらく私たちが好気性ゾーンで維持することを目的とした個々の心臓頻度の制御に関連していた。 しかし、BDNFの変化は身体活動、社会的相互作用および肯定的なストレスにも関連しており(Mattson、2008)、すべての研究が心血管訓練後にBDNFの増加を観察したわけで, 2014). 最後に、動物の研究は、協調が、持久力訓練ではなく、シナプス形成およびグリア変化を誘導することを示唆している(Black et al., 1990). 一緒に、BDNF分泌を駆動する名前の追加の要因は、フィットネス活動中よりもダンス中に重要であった可能性があり、したがって、ダンサーだけが最初の6月にBDNFの増加を示した理由を説明する。 BDNFレベルは、次の12ヶ月でベースラインに戻ったが、末梢血中のBDNFレベルに代表されるものよりも、成人の脳可塑性に関与する他の要因がなければならないことを示している。
運動誘発可塑性の神経生物学的メカニズムについても、脳予備の概念(Satz et al.,2011)が考慮されるべきである。 脳予備の概念は、異なるストレッサーに皮質回路を改造するためのより大きな動的能力を提供する増加したベースライン適応神経可塑性の個人差を, 2015).私たちの研究の結果は、長期的なダンス介入は、老化した人間の脳に神経可塑性を誘導する上で反復的な運動よりも優れていることを示唆している。
パースペクティブ
私たちの研究の結果は、 私たちは、この利点は、物理的、認知的、調整的な課題を組み合わせたダンスのマルチモーダルな性質に関連していると推測しています。 我々の知る限りでは、これは、高齢者の灰白質および認知低下を防止する手段としてダンスプログラムを推奨する最初の縦断的、無作為化研究である。 さらなる研究は、より詳細に時間的ダイナミクスとダンス誘発性神経可塑性の基礎となる神経生物学的メカニズムを明らかにするために必要とされ、この介入は本当にアルツハイマー病などの神経変性疾患のリスクを低減する可能性を秘めているかどうか。
著者の貢献
PMは、研究を設計し、実行し、データを分析し、記事を書きました。 KRは、研究を設計し、実行しました。 MSは、データを分析しました。 AHとVLは研究を設計しました。 MD:データ収集、記事の改訂。 TBとJKは、研究を設計し、データを分析しました。 NGMは研究を設計し、記事を書いた。
利益相反に関する声明
著者らは、この研究は、利益相反の可能性と解釈される可能性のある商業的または財政的関係がない場合に行われたと宣言している。
およびFriston,K. ボクセルベースの形態測定—方法。 11,805-821. 土井:10.1006/nimg.2000.0582
Pubmed Abstract|CrossRef Full Text|Google Scholar
Bamadis,P.D.,Vivas,A.B.,Styliadis,C.,Frantzidis,M.,Klados,M.,Schlee,W.,et al. (2014). 加齢における身体的および認知的介入のレビュー。 ニューロシ… バイオベハブ・バイオベハブ・バイオ 44巻206-220頁。 ドイ:10.1016/j.neubiorev.2014.03.019
PubMed Abstract|CrossRef Full Text|Google Scholar
Barulli,D.,And Stern,Y.(2013). 効率、容量、補償、維持、可塑性:cogniticeの予備の出現の概念。 トレンドCogn。 サイ… 17, 502–509. 土井:10.1016/j.tics.2013.08.012
Pubmed Abstract|CrossRef Full Text|Google Scholar
Black,J.E.,Isaacs,K.R.,Anderson,B.J.,Alcantara,A.A.,And Greenough,W.T.(1990). 学習はシナプス形成を引き起こし,運動活動は血管形成を引き起こし,成体ラットの小脳皮質では血管形成を引き起こす。 プロク… ナトル アカド サイ… 87,5568-5572 ドイ:10.1073/pnas.87.14.5568
PubMed Abstract|CrossRef Full Text|Google Scholar
Bliss,T.V.,And Lomo,T.(1973). 穿孔経路の刺激後のaneasthetizedウサギの歯状におけるシナプス伝達の長期的な増強。 J.フィジオール。 232, 331–356. 土井:10.1113/jphysiol.1973.sp010273
Pubmed Abstract|CrossRef Full Text|Google Scholar
Boyke,J.,Driemeyer,J.,Gaser,C.,Büchel,C.,May,A.(2008). 高齢者のトレーニング誘発性脳構造の変化。 J.Neurosci. 28, 7031–7035. 土井:10.1523/JNEUROSCI.0742-08.2008
PubMed Abstract|CrossRef Full Text|Google Scholar
Brown,S.,Martinez,M.,And Parsons,M.(2006). 人間のダンスの神経の基礎。 セレブ 第16代、1157年-1167年。 doi:10.1093/cercor/bhj057
PubMed Abstract|CrossRef Full Text/Google Scholar
Colcombe,S.J.,Erickson,K.I.,Scalf,P.E.,Kim,J.S.,Prakash,R.,McAuley,E.,et al. (2006). 有酸素運動の訓練は老化の人間の頭脳の容積を高める。 J.ジェロントール A.Biol。 サイ… メッド サイ… 61, 1166–1170. ドイ:10.1093/ジェロナ/61.11.1166
PubMed Abstract/CrossRef Full Text/Google Scholar
Echávarri,C.,Aalten,P.,Uyling,H.B.M.,Jacobs,H.I.L.,Visser,P.J.,Gronenschild,M.,et al. (2011). アルツハイマー病の初期のバイオマーカーとしての海馬傍回の萎縮。 脳の構造体。 ファンクト 215, 265–271. ドイ:10.1007/s00429-010-0283-8
PubMed Abstract|CrossRef Full Text|Google Scholar
Edelmann,E.,Lessmann,V.,And Brigadski,T.(2014). BDNF分泌におけるシナプス前および後のねじれおよびシナプス可塑性における作用。 神経薬理学76,610-627. ドイ:10.1016/j.neuropharm.2013.05.043
Pubmed Abstract|CrossRef Full Text|Google Scholar
Erickson,K.I.,Voss,M.W.,Prakash,R.S.,Basak,C.,Szabo,A.,Chaddock,l.,et al. (2011). 運動訓練は海馬の大きさを増加させ、記憶を改善する。 プロク… ナトル アカド サイ… 108,3017-3022. ドイ:10.1073/pnas.1015950108
PubMed Abstract|CrossRef Full Text|Google Scholar
Erickson,K.I.,Weinstein,A.M.,and Lopez,O.L.(2012). 身体活動、脳の可塑性、およびアルツハイマー病。 アーチ メッド Res.43,615-621. 土井:10.1016/j.arcmed.2012.09.008
Pubmed Abstract|CrossRef Full Text|Google Scholar
Flöel,A.,Ruscheweyh,R.,Krüger,K.,Willemer,C.,Winter,B.,Völker,K.,et al. (2010). 身体活動および記憶機能:neurotrophinsおよび大脳の灰白質の容積は行方不明のリンクであるか。 49,2756-2763 土井:10.1016/j.neuroimage.2009.10.043
Pubmed Abstract|CrossRef Full Text|Google Scholar
Freret,T.,Gaudreau,P.,Schumann-Bard,P.,Billard,J.M.,Popa-Wagner,A.(2015). 後期うつ病に対する脳予備の神経保護効果の根底にあるメカニズム。 J.ニューラル トランスム (ウィーン)122、S55-S61。 ドイ:10.1007/s00702-013-1154-2
PubMed Abstract|CrossRef Full Text/Google Scholar
Gregory,S.M.,Parker,B.,And Thompson,P.D.(2012). 身体活動、認知機能、および脳の健康:認知症の予防における運動訓練の役割は何ですか? 脳科学 2, 684–708. doi:10.3390/brainsci2040684
Pubmed Abstract|CrossRef Full Text|Google Scholar
Hänggi,J.,Koeneke,S. ることを示した。 専門の女性のバレエダンサーのsensorimotorネットワークの構造neuroplasticity。 ハム ブレインマップ 31, 1196–1206. ドイ:10.1002/hbm.20928
PubMed Abstract|CrossRef Full Text|Google Scholar
Helmstaedter,C.,And Durwen,H.F.(1990). 言語学習と保持テスト。 口頭記憶性能を評価するのに便利で差別化されたツール。 シュヴァイツシュヴァイツシュヴァイ アーチ ニューロール 精神科医。 (1985) 141, 21–30. PubMed Abstract/Google Scholar
Huang,E.J.,And Reichardt,L.F.(2001). Neurotrophins:神経の開発および機能における役割。 アンヌ ニューロシー牧師。 24, 677–736. ドイ:10.1146/annurev.ニューロ24.1.677
PubMed Abstract|CrossRef Full Text|Google Scholar
Hüfner,K.,Binetti,C.,Hamilton,D.A.,Stephan,T.,Flanagin,V.L.,Linn,J.,et al. (2011). プロのダンサーやスラックライナーにおける海馬形成の構造的および機能的可塑性。 海馬21、855-865。 土井:10.1002/hipo.20801
PubMed Abstract|CrossRef Full Text|Google Scholar
Karvonen,M.J.,Kentala,E.,And Mustala,O. (1957). トレーニングが心拍数に及ぼす影響:縦断研究。 アン メッド 経験値 バイオル フェン 35, 307–315. PubMed Abstract/Google Scholar
Kattenstroth,J.C.,Kalisch,T.,Holt,S.,Tegethoff,M.,And Dinse,H.R.(2013). ダンス介入の半年は、心臓呼吸機能に影響を与えることなく、高齢者の姿勢、感覚運動、および認知能力を向上させます。 フロント 加齢性神経疾患。 5:5. ドイ:10.3389/fnagi.2013.00005
PubMed Abstract|CrossRef Full Text|Google Scholar
Kattenstroth,J.C.,Kolankowska,I.,Kalisch,T. およびDinse,H.R.(2010)。 複数年のダンス活動を持つ高齢者における優れた感覚、運動、および認知能力。 フロント 加齢性神経疾患。 2:31. ドイ:10.3389/fnagi.2010.00031
PubMed Abstract|CrossRef Full Text|Google Scholar
Kempermann,G.,Kuhn,H.G.,and Gage,F.H.(1997). 豊かな環境に住んでいる大人のマウスのより多くの海馬ニューロン。 自然。 386, 493–495. doi:10.1038/386493a0
PubMed Abstract|CrossRef Full Text|Google Scholar
Kirk-Sanchez,N.J.,And McGough,E.L.(2014). 高齢者における身体運動と認知能力:現在の視点。 クリン インターヴ 年9月51-62日… 土井:10.2147/CIA.S39506
Pubmed Abstract|CrossRef Full Text|Google Scholar
Lessmann,V.,And Brigadski,T.(2009). シナプスBDNF分泌のメカニズム、位置、および速度論:更新。 ニューロシ… 昭和65年11月22日に廃止された。 土井:10.1016/j.neures.2009.06.004
Pubmed Abstract|CrossRef Full Text|Google Scholar
Maass,A.,Düzel,S.,Goerke,M.,Becke,A.,Sobieray,U.,Neumann,K.,et al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M. ら,hokelmann,A.,Kaufmann,J.,et a l. (2016). 健康な先輩の灰色の頭脳の問題に対するダンスおよび健康のスポーツの訓練の効果。 スポーツウィス 46, 213-222. ドイ:10.1007/s12662-016-0411-6
CrossRef全文/Google Scholar
Niemann,C.,Godde,B.,And Voelcker-Rehage,C.(2014). 心血管だけでなく、協調運動も高齢者の海馬容積を増加させる。 フロント 加齢性神経疾患。 6:170. ドイ:10.3389/fnagi.2014.00170
PubMed Abstract|CrossRef Full Text/Google Scholar
Oswald,W.D.,Gunzelmann,T. ら、rupprecht,R.,およびHagen,B.,(2 0 0 6)。 高齢者との単一対複合認知および身体訓練の差動効果:5年の視点でのSimA研究。 ユーロ J.エイジング3,179-192. ドイ:10.1007/10433-006-0035-z
CrossRef全文/Google Scholar
Pantel,J.,Kratz,B.,Essig,M.,And Schröde,J.(2003). 加齢に伴う認知機能低下を有する被験者における海馬傍容積欠損。 アム… J.精神医学160,379-382. 土井:10.1176/安比.ajp。160.2.379
PubMed Abstract|CrossRef全文|Google Scholar
Park,H.、およびPoo、M.M.(2013)。 神経回路の開発および機能のNeurotrophinの規則。 ナット ニューロシー牧師。 14, 7–23. doi:10.1038/nrn3379
Pubmed Abstract|CrossRef Full Text|Google Scholar
Rehfeld,K.,Hökelmann,A.,Kaufmann,J.,And Müller,N.G.(2016). “健康なsenioresの頭脳の可塑性、バランスの性能および注意に対するダンス対適性の訓練の効果:pairwise縦方向のグループの比較のためのSPM12の新しいアプローチ、”Proceeding-International Conference Of Sports and Neuroscience(Magdeburg)、127-138。
Satz,P.,Cole,M.A.,Hardy,D.J.,And Rassovsky,Y.(2011). 脳と認知予備:メディエーター(複数可)と妥当性、批判を構築します。 J.Clin. 経験値 ニューロサイチョル… 33, 121–130. doi:10.1080/13803395.2010.493151
PubMed Abstract|CrossRef Full Text|Google Scholar
Schega,L.,Peter,B.,Brigadski,T.,Leşmann,V.,Isermann,B.,Hamacher,D.,et al. (2016). 高齢者における好気性能力および認知機能に対する間欠的な正常圧低酸素症の影響。 J.Sci. メッド スポーツ19,941-945. 土井:10.1016/j.jsams.2016.02.012
Pubmed Abstract|CrossRef Full Text|Google Scholar
Taube,W.(2008). バランストレーニングに応答した神経生理学的適応。 Dtsch。 Z.Sportmed. 63, 273–277. ドイ:10.5960/dzsm.2012.030
CrossRef全文|Google Scholar
Taubert,M.,Draganski,B.,Anwander,A.,Müller,K.,Horstmann,A.,Villringer,a.,et al. (2010). 人間の脳構造の動的特性:皮質領域および関連する繊維接続における関連する変化を学習する。 J.Neurosci. 30, 11670–11677. 土井:10.1523/JNEUROSCI.2567-10.2010
PubMed Abstract/CrossRef Full Text/Google Scholar
Thomas,A.G.,Dennis,A.,Bandettini,P.A.,And Johansen-Berg,H.(2012). 脳の構造に及ぼす好気性活性の影響。 フロント サイコル 3:86. ドイ:10.3389/fpsyg.2012.00086
Pubmed Abstract|CrossRef Full Text|Google Scholar
Tisserand,D.J.,Pruessner,J.C.,Sanz Arigita,E.J.,van Boxtel,M.P.,Evans,A.C.,Jolles,J.,et al. (2002). 地域の前頭皮質のボリュームは、加齢に差動的に減少します: 容積測定アプローチとボクセルベースの形態測定を比較するためのMRI研究。 17,657-669. doi:10.1016/s1053-8119(02)91173-0
Pubmed Abstract|CrossRef Full Text/Google Scholar
van Praag,H.,Shubert,T.,Zhao,C.,and Gage,F.H.(2005). 運動は、高齢マウスの学習と海馬神経新生を強化します。 J.Neurosci. 25, 8680–8685. 土井:10.1523/JNEUROSCI.1731-05.2005
PubMed Abstract|CrossRef Full Text|Google Scholar
Verghese,J.,Lipton,R.B.,Katz,M.J.,Hall,C.B.,Dervy,C.A.,Kuslansky,G.,et al. (2003). レジャー活動と高齢者の認知症のリスク。 N.Engl. J.Med. 348, 2508–2516. doi:10.1056/Nejmoa022252
Pubmed Abstract|CrossRef Full Text/Google Scholar
Vital,T.M.,Stein,A.M.,de Melo Coelho,F.G.,Arantes,F.J.,Teodorov,E.,Santos-Galduróz,R.F.,et al. (2014). 高齢者における身体運動および血管内皮増殖因子(VEGF):系統的レビュー。 アーチ ジェロントール ジェリアトル 59, 234–239. 土井:10.1016/j.archger.2014.04.011
PubMed要約|CrossRef全文|Google Scholar
Voelcker-Rehage,C. ることを示した。 構造的および機能的な脳の変化は、寿命にわたるさまざまな種類の身体活動に関連しています。 ニューロシ… バイオベハブ・バイオベハブ・バイオ 37,2268-2295 ドイ:10.1016/j.neubiorev.2013.01.028
PubMed Abstract|CrossRef Full Text|Google Scholar
Zatorre,R.J.,Fields,R.D.,And Johansen-Berg,H.(2012). 灰色と白の可塑性:学習中の脳構造の神経イメージング変化。 ナット ニューロシ… 15, 528–536. ドイ:10.1038/nn.3045
PubMed Abstract|CrossRef Full Text/Google Scholar
Zimmermann,P.,and Fimm,B.(2002). 注意テスト(タップ)、バージョン1.7、マニュアル—Part1のためのテストバッテリー。 ヴュルセレン:Psytest。
div