
Le carcinome épidermoïde (CSC), également connu sous le nom de carcinome à cellules épineuses, est l’une des principales causes de décès liés au cancer dans le monde. Le CSC est une tumeur épithéliale maligne, apparaissant dans les tissus qui constituent une barrière entre un organisme et l’environnement, tels que la peau, la bouche, la cavité, l’œsophage et le poumon. À un stade précoce, le CSC s’accompagne d’une kératinisation épidermique et d’une formation d’ulcère à la surface de la muqueuse, et les tissus plus profonds sont envahis par le CSC à un stade ultérieur. Pour leur aspect microscopique, les cellules CSC sont disposées en nids, qui sont entourés de cellules polygonales avec des bordures cellulaires distinctes et des noyaux hyperchromatiques. Les SCCS ont une forte tendance à métastaser, généralement par les ganglions lymphatiques régionaux, et peuvent causer des dommages systématiques à plusieurs organes. La chirurgie, la thérapie au laser et la radiothérapie continuent d’être le traitement le plus utilisé pour les CSC. Les caractéristiques cytomorphologiques uniques des variantes du CSC conduisent à un traitement et à des résultats distincts.
L’antigène de surface cellulaire CD109 est une glycoprotéine liée au glycosylphosphatidylinositol (GPI) d’environ 170 kDa et un membre de la famille des macroglobulines a2 (a2M)/C3, C4, C5 de protéines contenant du thioester. Le gène CD109 humain est situé sur le chromosome 6q, constituant environ 3,3% de la séquence génomique CD109 totale avec ses 33 exons. L’adnc CD109 isolé comprend une trame de lecture ouverte de 4335 pb codant pour un acide aminé 1445 (aa). La protéine CD109 contient un peptide leader N-terminal de 21 aa, une région d’appât putative (aa 651-683), un site de liaison au thioester (aa 918-924), un hexapeptide définissant la réactivité du thioester (aa1030–1035) et une séquence de signal d’ancrage GPI de consensus C-terminal avec le clivage prévu après l’acide aminé 1420 (Fig. 1) . Le peptide leader N-terminal ancre la protéine à la membrane interne dans le périplasme et le fragment N-terminal de CD109 sécrété par les cellules après clivage par la protéase de la furine. Le clivage protéase d’une région d’appât largement désordonnée active CD109, entraînant un changement conformationnel qui emprisonne la protéase dans une structure en forme de cage et expose la liaison thioester hautement réactive. Le site de liaison au thioester qui, lors de l’activation, peut lier de manière covalente CD109 aux résidus de lysine à la surface de la protéase attaquante. En outre, la séquence de signaux d’ancrage GPI de consensus C-terminal est également connue sous le nom de domaine de liaison aux récepteurs.

The structure of CD109 protein. La pleine longueur de la protéine CD109 comprend 1445 acides aminés (aa), y compris un peptide signal N-terminal de 21 aa, une région d’appât putative (aa 651-683), un site de liaison au thioester (aa 918-924), un hexapeptide définissant la réactivité du thioester (aa1030–1035) et une séquence de signal d’ancrage GPI de consensus C-terminal (aa 1420-1445) avec le clivage prévu pour se produire après l’acide aminé 1420
CD109 a d’abord été identifié comme un antigène de surface cellulaire par un anticorps monoclonal élevé contre la lignée cellulaire primitive lymphoïde/myéloïde KG1a. Des études antérieures ont rapporté que CD109 est exprimé sur un sous-ensemble de cellules mononucléées de moelle osseuse positives CD34 + fœtales et adultes, de lymphoblastes T activés, de plaquettes activées, de cellules endothéliales, de sous-ensembles de cellules souches mésenchymateuses et de plusieurs lignées de cellules tumorales humaines, mais pas exprimé dans les cellules T humaines au repos, les plaquettes ou les leucocytes du sang périphérique. En particulier, le CD109 a été détecté dans les CSC de plusieurs organes, notamment le poumon, l’œsophage, le col utérin, la peau, le pénis, la cavité buccale et la vésicule biliaire. CD109 est un composant du système de récepteur du facteur de croissance transformant-β1 (TGF-β1) et régule négativement la signalisation du TGF-β1. Des études sur la perte et le gain de fonctions ont suggéré que CD109 pourrait être une cible moléculaire pour le développement de nouveaux traitements pour les CSC d’origines tissulaires diverses. Dans cette revue, nous présenterons un résumé des connaissances actuelles sur le CD109 et sa relation avec les CSC.
Expression de CD109 dans les SCCs
Dans les tissus tumoraux, CD109 a été détecté immunohistochimiquement dans les SCCs ainsi que dans les carcinomes urothéliaux, les mélanomes malins, les carcinomes mammaires basaux, le myxofibrosarcome, les sarcomes épithéliaux et le gliome. En particulier, Shiraki et al. des cellules tumorales périvasculaires positives au CD109 rapportées dans des tissus de gliome humain de grade inférieur et dans un modèle murin ont récapitulé le gliome humain, suggérant un rôle clé du CD109 pour cette maladie. Des études antérieures ont montré que l’expression élevée de CD109 dans les CSC et l’expression limitée dans les cellules squameuses normales (tableau 1). De plus, CD109 est fortement exprimé dans des CSC bien différenciés plutôt que dans des CSC modérément ou mal différenciés, ainsi le niveau d’expression de CD109 est inversement corrélé au grade tumoral.
CD109 et CSC pulmonaire
Le carcinome épidermoïde pulmonaire (LSCC) est l’un des principaux sous-types de carcinomes non épidermoïdes. – cancer du poumon à petites cellules (CPNPC), et représente environ 20 à 30% des cas de CPNPC. L’expression de CD109 a été examinée dans les carcinomes à cellules pulmonaires humaines par RT-PCR quantitative, qui a montré une expression significativement plus élevée de CD109 dans les carcinomes épidermoïdes, mais pas dans les adénocarcinomes, les carcinomes à grandes cellules ou les carcinomes à petites cellules. De plus, Sato et coll. a utilisé un anticorps CD109 pour analyser l’expression de CD109 dans les tissus normaux du poumon et du carcinome pulmonaire. Dans les tissus pulmonaires normaux, l’expression du CD109 se limitait principalement aux cellules basales de l’épithélium bronchique et bronchiolaire. Dans les tissus du carcinome pulmonaire, une forte immunoréactivité a été détectée préférentiellement dans les LSCC, mais pas dans les autres types de carcinomes pulmonaires. Ces résultats indiquent que le CD109 est spécifiquement réglementé à la hausse dans les LSCC.
Le CD109 et le CSC de l’œsophage
Le carcinome épidermoïde de l’œsophage (ESCC) représente un sixième de toute la mortalité liée au cancer, avec une incidence plus élevée dans les pays asiatiques. Le pronostic de l’ESCC reste médiocre et le taux de survie à 5 ans de l’ESCC est d’environ 14 à 22%. Des études récentes suggèrent qu’une expression altérée des récepteurs TGF-β contribue à la progression de l’ESCC, et qu’une expression élevée des protéines inhibitrices de la signalisation du TGF-β est corrélée à un mauvais pronostic de l’ESCC. Hashimoto et coll. on a examiné l’expression du CD109 dans les tissus de l’ESCC par RT-PCR quantitative, et la transcription de l’ARNm du CD109 a été régulée à la hausse dans environ 50 % des ESCC. Plus tard, le taux de protéines de CD109 dans l’ESCC a été examiné par immunohistochimie sur des puces tissulaires (TMA). Dans le tissu oesophagien non malade, l’expression de CD109 a été limitée dans le cytosol des cellules épithéliales stratifiées à un niveau faible. Dans tous les échantillons d’ESCC provenant des AMT, des brins de cellules épithéliales malignes malignes ont montré une forte coloration CD109 dans le cytosol. Aucune coloration positive au CD109 n’a été observée chez d’autres types de cellules. De plus, l’expression de CD109 était plus élevée dans les CCE bien et modérément différenciées que dans les CCE peu différenciées.
CD109 et SCC du col utérin
Le cancer du col utérin (col utérin) représente environ 12% des décès liés au cancer des femmes. Les CSC représentent 80 à 85% de tous les cancers du col de l’utérus. Zhang et coll. a montré que l’expression de CD109 était significativement plus élevée dans les CSC cervicaux que dans les adénocarcinomes de l’endomètre. Ils ont également étudié l’expression de CD109 dans cinq lignées de cellules de carcinome cervical humain et observé des niveaux élevés d’expression de CD109 dans deux lignées de cellules SCC.
CD109 et CSC cutané
Le carcinome épidermoïde cutané (CSCC) est le deuxième type de cancer de la peau non mélanome (NMSC) le plus fréquent avec une incidence en augmentation constante. Bien que les CSCC aient un pronostic généralement favorable, il y a encore environ 1,5 à 2% des patients qui meurent de cette maladie. L’expression du CD109 dans les CSCCs a été examinée par immunohistochimie sur les AMT. Dans les peaux normales, CD109 était faiblement exprimé dans la couche basale des cellules épidermiques, tandis que des brins de cellules épidermiques malignes malignes présentaient une forte coloration CD109. En outre, l’expression de CD109 était inversement corrélée avec les grades CSCC. Comme les CSC, l’expression de CD109 était plus élevée chez les CSCC bien et modérément différenciées que chez les CSCC peu différenciées. Le carcinome épidermoïde du pénis (CSCC) est un sous-type de CSCC et a rarement été étudié. Dong et coll. démontré que la protéine CD109 est fortement exprimée dans les cellules malignes squameuses des PSCC par rapport aux tissus péniens normaux sur les AMT. De plus, le motif d’expression est validé sur des échantillons de CPCC chirurgicaux frais par immunofluorescence, qRT-PCR et western blot, suggérant que CD109 peut être un biomarqueur pour la CPCC.
Le CD109 et le CSC de la cavité buccale
Le carcinome épidermoïde buccal (OSCC) représente 2 à 3 % de tous les cancers dans le monde. La perte de mutation fonctionnelle dans le récepteur TGF-β de type II est un événement fréquent pour le CSC de la cavité buccale. L’expression du CD109 dans les tissus buccaux normaux et les tissus OSCC de 124 patients a été examinée par coloration immunohistochimique. Des niveaux élevés d’expression de CD109 ont été fréquemment détectés dans les CSC et les lésions prémalignées de la cavité buccale, mais pas dans les épithéliums squameux normaux. De plus, le niveau d’expression de CD109 était significativement plus élevé dans les OSCC bien différenciées que dans les OSCC modérément ou mal différenciées, ce qui implique que l’expression de CD109 est corrélée aux stades de différenciation des OSCC. De plus, les lignées cellulaires OSCC surexprimant CD109 ont montré une croissance cellulaire accélérée in vitro, ce qui implique que CD109 participe à la progression des OSCC.
CD109 et CSC de la vésicule biliaire
Le cancer de la vésicule biliaire (CBG) est le plus agressif des cancers biliaires dont la survie médiane est la plus courte. Le carcinome épidermoïde de la vésicule biliaire (GSCC) représente 3% du néoplasme malin de cet organe. Des sous-types de tissus GBCs, y compris l’adénocarcinome (AC), le carcinome épidermoïde (SCC) et le carcinome adénosquameux (ASC), ont été examinés sur des AMT par coloration immunohistochimique avec un anticorps CD109. La coloration au CD109 était négative dans tous les tissus normaux de la vésicule biliaire et des tissus AC. Pendant ce temps, des cellules CD109 positives ont été trouvées dans 86,7% des CSC et 91,7% des ASC. Comme CD109 est clairement exprimé dans les cellules sqameuses malignes de la vésicule biliaire, CD109 peut être un marqueur diagnostique des CSC et des ASC de la vésicule biliaire.
Voie de signalisation TGF-β et SCCs
La voie de signalisation TGF-β est impliquée dans de nombreux processus cellulaires, notamment la croissance cellulaire, l’apoptose de différenciation cellulaire et l’homéostasie cellulaire. La famille des ligands TGF-β, TGF-β1, TGF-β2 et TGF-β3, se lie à des récepteurs transmembranaires spécifiques de sérine/thréonine kinase de type I et de type II (TGF-ßR1 et TGF-ßR2), ce qui entraîne l’activation de l’activité de la kinase TGF-ßR1. Le TGF-ßR1 activé propage ensuite le signal en phosphorylant ses substrats intracellulaires, les R-SMADs (Smad2 et Smad3). Smad2 et Smad3 interagissent avec TGF-ßR1 et SARA (Smad anchor for receptor activation), une protéine du domaine FYVE qui interagit directement avec Smad2 et Smad3, SARA fonctionne pour recruter Smad2 sur le récepteur TGF-β, puis les R-SMADs phosphorylés forment des complexes hétéromères avec Co-SMAD (Smad4). Après la phosphorylation et les complexes ultérieurs avec Smad4, ces complexes R-Smads sont libérés du TGF-ßR1 et du SARA, puis se translocent dans le noyau où ils interagissent avec des facteurs de transcription qui les recrutent en éléments promoteurs spécifiques des gènes cibles.
L’endocytose des récepteurs est un mécanisme de régulation pivot dans la transduction du signal. Les récepteurs TGF-β sont internalisés par des voies dépendantes de la clathrine et des cavéoles. L’internalisation des récepteurs TGF-β via les fosses revêtues de clathrine a été liée à la signalisation via Smad2 / 3 et au recyclage des récepteurs. En revanche, la localisation du récepteur TGF-β dans les cavéoles est associée à une régulation négative de la signalisation Smad2 / 3 et à une dégradation du récepteur suite à l’ubiquitination par la ligase E3-ubiquitine Schtroumpf2. Cependant, les Smad inhibiteurs (Smad6 et Smad7) forment une sous-classe distincte de Smad qui agissent de manière opposée aux R-Smad et antagonisent la signalisation. Ils peuvent rivaliser avec les R-Smads pour se lier au TGF-ßR1 activé et inhiber ainsi la phosphorylation des R-Smads. De plus, ils recrutent des ligases E3-ubiquitine sur le TGF-ßR1 activé, ce qui entraîne une ubiquitination et une dégradation des récepteurs et la fin de la signalisation.
La dérégulation de la voie TGF-β a été impliquée dans plusieurs types de cancer. Des études ont démontré que la signalisation du TGF-β provoque un effet préventif pendant les premiers stades de la tumorigenèse, mais un effet suppresseur pendant les derniers stades du développement tumoral. Des mutations du gène TGF-ßR1 ont également été trouvées dans les CCS de la peau, suggérant que l’inactivation du TGF-β conduit à l’initiation des CCS.
Voie de signalisation CD109 et TGF-β
CD109 est un co-récepteur TGF-β et module l’activité du récepteur de signalisation TGF-β de manière spécifique à la cellule. À la surface de la cellule, CD109 régule négativement la voie de signalisation du TGF-β1 via la formation d’un complexe récepteur avec le TGF-ßR1 et le TGF-ßR2 dans les kératinocytes humains. Les récepteurs TGF-β se dégradent après l’ubiquitination par la ligase E3-ubiquitine Schtroumpf2 et sont internalisés par des voies dépendantes de la clathrine et des cavéoles. Bizet et coll. a démontré que CD109 s’associe à la cavéoline-1 et favorise l’endocytose des récepteurs TGF-β. De plus, CD109 favorise la localisation des récepteurs TGF-β dans le compartiment cavéolaire en présence de ligand et facilite la dégradation des récepteurs TGF-β. CD109 régule également la localisation et l’association de Smad7/ Schtroumpf2 avec TGF-ßR1. Les effets inhibiteurs de CD109 nécessitent l’expression de Smad7 et l’activité de l’ubiquitine ligase de Schtroumpf2. De plus, CD109 peut être libéré de la surface cellulaire par des lipases cellulaires telles que la phospholipase C spécifique au phosphatidylinositol (PI-PLC). La forme soluble de CD109 conserve sa capacité à se lier au TGF-β1 et à le confisquer des récepteurs du TGF-β.
Cependant, Vorstenbosch et al. rapporté que CD109 a régulé de manière différentielle les voies ALK1-Smad1 / 5 induites par le TGF-β par rapport aux voies ALK5-Smad2 / 3 (ALK1 et ALK5 sont tous des récepteurs TGF-β de type I). Ils ont découvert que la signalisation TGF-β inhibe la prolifération et la migration des cellules endothéliales, tandis que la signalisation TGF-β induit également ces processus via ALK1-Smad1 /5. Ils démontrent que ALK1 est exprimé et co-localise avec CD109 dans les kératinocytes de souris et que les souris surexprimant CD109 dans l’épiderme présentent une signalisation ALC1-Smad1/5 améliorée, mais une signalisation ALC5-Smad2/3 diminuée.
En outre, le TGF-β1 est un inhibiteur puissant de la croissance dans la plupart des cellules épithéliales. Hagiwara et al. a démontré que la surexpression orale des lignées cellulaires du CSC CD109 accélérait la prolifération cellulaire et altérait l’effet anti-prolifératif médié par le TGF-β1. En revanche, les cellules SCC avec un knockdown CD109 ont montré une croissance cellulaire plus lente. Un niveau élevé d’expression de CD109 a inhibé la phosphorylation de Smad2, atténuant ainsi la signalisation TGF-β1 / Smad2 et altérant la suppression de la croissance cellulaire médiée par TGF-β1, le knockdown CD109 a augmenté la phosphorylation de Smad2 par stimulation de TGF-β1. Bien que CD109 régule également la signalisation Smad1 / 5, il n’a pas été lié au développement de SCC. Ensemble, CD109 facilite le développement de SCCs via l’inhibition de la voie TGF-β-Smad2/3 (Fig. 2).

Signalisation CD109 et TGF-β. Le TGF-β lie le TGF-ßR2, qui recrute et phosphoryle le TGF-ßR1. Le TGF-ßR1 activé propage le signal en aval en phosphorylant directement Smad2 et Smad3. Les R-SMADs phosphorylés forment ensuite des complexes hétéromères avec Smad4, combinés à des facteurs de transcription, régulent la transcription des gènes et la fonction cellulaire. Les récepteurs TGF-β s’intériorisent via les fosses recouvertes de clathrine ou la voie cavéolaire. CD109 augmente la liaison du TGF-β aux récepteurs du TGF-β et favorise la localisation du récepteur du TGF-β dans les cavéoles, augmente finalement l’endocytose du récepteur du TGF-β et facilite la dégradation du récepteur du TGF-β. Les effets inhibiteurs de CD109 nécessitent l’expression de Smad7 et l’activité de l’ubiquitine ligase de Smurf2
Signalisation CD109 et STAT3
Le transducteur de signal et l’activateur du facteur de transcription 3 (STAT3) sont essentiels à la prolifération, à la différenciation, à la migration, à la survie et à l’oncogenèse des cellules. Litvinov et coll. a rapporté que l’expression de la protéine CD109 était nettement diminuée dans l’épiderme psoriasique par rapport à la peau adjacente non impliquée. Cependant, l’expression de l’ARNm CD109 est inchangée dans les plaques psoriasiques par rapport à la peau normale, suggérant une possibilité que la libération de la protéine CD109 soit améliorée dans les kératinocytes psoriasiques. Ils ont suggéré que le CD109 libéré / soluble est capable d’induire des changements moléculaires connus dans le psoriasis. In vitro, ils ont constaté que la transfection de l’ARNsi CD109 régule la STAT3, la libération de CD109 à partir de la surface cellulaire des kératinocytes humains en culture. De plus, le CD109 exogène / recombinant induit une signalisation STAT3 dans les kératinocytes humains. En outre, Chuang et al. l’expression de CD109 a été fortement régulée dans les cellules d’adénocarcinome pulmonaire métastatique, et les cellules exprimant un shRNA CD109 (shCD109) ont montré une réduction spectaculaire de la phosphorylation de STAT3. Le knockdown STAT3 a considérablement réduit les métastases et la restauration de l’activité STAT3 a augmenté la capacité des cellules exprimant le shCD109 à métastaser. Lors de l’activation, STAT3 est phosphorylée par la protéine non réceptrice tyrosine kinases janus kinase 2 (JAK2), conduisant à la formation du dimère STAT3 et à la translocation dans le noyau. Cependant, l’inhibition de l’activité de la JAK kinase dans les fibroblastes surexprimant CD109 a réduit la STAT3 phosphorylée à un niveau similaire à celui des cellules parentales exprimant de faibles niveaux de CD109, suggérant que la phosphorylation de la STAT3 induite par CD109 nécessite une activité de la JAK kinase. Ainsi, la signalisation JAK / STAT3 pourrait médier les effets du CD109 sur la croissance tumorale et les métastases.
Bien que la suppression du CD109 dans les kératinocytes humains et les cellules de l’adénocarcinome pulmonaire régule à la baisse la signalisation de STAT3 in vitro, les souris déficientes en CD109 ont montré des résultats opposés. Mii et coll. souris déficientes en CD109, qui présentaient des anomalies cutanées, y compris une hyperplasie épithéliale, et une infiltration cellulaire inflammatoire. Ils ont rapporté que la phosphorylation de STAT3 chez les souris déficientes en CD109 était significativement plus élevée que chez les souris de type sauvage. De plus, la régulation ascendante de la signalisation STAT3 est associée à une prolifération accrue et à une différenciation altérée des kératinocytes.
La divergence des résultats des études in vitro et in vivo pourrait être causée par les changements systématiques du microenvironnement dans les tissus de souris déficientes en CD109. La perte de CD109 dans toutes les cellules de la souris pourrait modifier le microenvironnement sous-cutané qui active la signalisation STAT3 dans les kératinocytes. De plus, CD109 peut exercer des effets régulateurs distincts dans différents types de cellules, conduisant à une modification spécifique du type de cellule dans la signalisation STAT3. En plus des kératinocytes, CD109 est exprimé dans les cellules endothéliales, les cellules épithéliales et les fibroblastes, qui participent à la constitution du tissu cutané. Cependant, à ce jour, la relation entre la signalisation CD109 et STAT3 n’a pas été explorée dans ces types de cellules.
Signalisation CD109 et EGFR
Le récepteur du facteur de croissance épidermique (EGFR) est un membre de la famille des récepteurs ErbB. Lors de la liaison du ligand par l’EGF, l’EGFR forme des dimères, homodimères ou hétérodimères avec un autre membre de la famille ERBB HER2. Les récepteurs dimérisés s’auto-phosphorylent, puis phosphorylent la protéine non réceptrice tyrosine c-Src kinase, qui active STAT3. L’activation de l’EGFR favorise la migration, la survie et la prolifération des cellules. Dans les tumeurs malignes, la surexpression de l’EGFR est corrélée à la profondeur d’invasion de la tumeur et liée à un pronostic plus défavorable. Les mutations qui conduisent à une surexpression de l’EGFR sont détectées dans le CSC pulmonaire, le CSC de la tête et du cou et le CSC de l’œsophage. Le CD109 ancré à la membrane dans les cellules SK-MG-1 interagit directement avec l’EGFR et améliore la signalisation de l’EGF, ce qui augmente ensuite la migration et l’invasion cellulaires, tandis que le CD109 sécrété n’a aucun effet sur la signalisation de l’EGF. L’EGFR pourrait médier les effets du CD109 sur la signalisation STAT3, ce qui nécessite d’autres études pour élucider (Fig. 3).
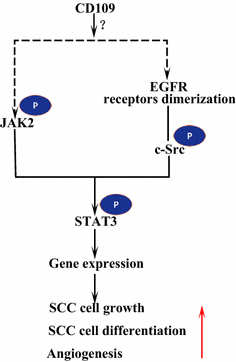
Signalisation CD109 et EGFR/STAT3. CD109 facilite la signalisation EGFR. Les récepteurs EGFR dimérisés phosphorylent c-Src, qui à son tour phosphorylent STAT3. CD109 peut également activer STAT3 par phosphorylation de JAK2, puis favoriser la croissance cellulaire du CSC, la différenciation et l’angiogenèse
À ce jour, le tableau général de la tumorigénèse et du développement du CSC reste incomplet. Il y a des discussions croisées entre les voies de signalisation TGF-β et STAT3 ou EGFR. CD109 est probablement l’un des effecteurs clés du réseau de signalisation régulant les SCC. Cependant, aucune preuve directe n’a été rapportée pour délimiter les rôles du CD109 dans la pathogenèse des CSC.
Perspectives
Des études à partir d’échantillons de tissus humains indiquent que le CD109 est fortement exprimé dans les CSC de plusieurs organes, en particulier dans les cellules malignes malignes bien différenciées. En détectant l’expression de CD109 par immunohistochimie dans les tissus humains, CD109 peut potentiellement agir comme un biomarqueur pour déterminer la progression du CSC. Les études actuelles suggèrent que CD109 est fortement exprimé dans les CSC bien différenciés et son expression est plus faible dans les CSC non différenciés. Cependant, il n’est pas clair si CD109 est associé à une invasion vasculaire, à des métastases et à un pronostic après une intervention chirurgicale. Par conséquent, d’autres études sont nécessaires pour explorer la signification clinicopathologique du CD109 dans les CSC dans un échantillon de plus grande taille.
CD109 est une glycoprotéine liée au GPI, qui lui permet d’être libérée de la membrane. La forme soluble de CD109 affecte également la liaison du TGF-β à ses récepteurs et module ensuite la progression du CSC. Litvinov et coll. a constaté que CD109 libéré de la surface cellulaire dans le milieu extracellulaire, et la forme libérée de CD109 conserve sa capacité à induire des voies de signalisation intracellulaires. En outre, Sakakura et al. rapporté que le sérum CD109 a été libéré par une tumeur xénogreffée et qu’il augmente proportionnellement au volume de xénogreffe tumorale. Par conséquent, la détection du taux sérique de CD109 pourrait aider à surveiller les tumeurs surexprimant CD109, y compris les CSC.
Les exosomes communiquent la lésion tumorale primaire et sa niche via son emballage contenant des protéines sélectionnées ou d’autres molécules. L’isolement et les analyses de l’exosome tumoral circulant peuvent servir de biomarqueurs pour le diagnostic des patients cancéreux. Le CD109 exogène a été identifié comme un composant de l’exosome sécrété à partir de 293 cellules transfectées, ce qui en fait une cible prometteuse pour le diagnostic des exosomes. Pourtant, à ce jour, CD109 n’a pas été rapporté dans les exosomes dérivés du CSC ou d’autres cellules tumorales. Les mécanismes sous-jacents de l’empaquetage du CD109 dans les exosomes méritent une étude plus détaillée.
En plus de son potentiel en tant que biomarqueur, le CD109 pourrait être une cible pour les approches thérapeutiques. CD109 est une protéine membranaire, qui peut être ciblée directement par des anticorps ou des enzymes spécifiques. CD109 pourrait également être reconnu par des véhicules ciblés pour l’administration de médicaments. Cependant, les rôles détaillés du CD109 dans la pathogenèse des CSC ne sont toujours pas clairs. Par exemple, la relation d’expression inférieure de CD109 et de CSC indifférenciés doit être définie. L’intervention contre le CD109 ne peut être envisagée en pratique clinique qu’après une évaluation minutieuse des risques et des bénéfices.