
carcinomul cu celule scuamoase (SCC), cunoscut și sub numele de carcinom cu celule înțepătoare, este una dintre principalele cauze ale decesului legat de cancer la nivel mondial. SCC este o tumoare epitelială malignă, care apare în țesuturi care asigură o barieră între un organism și mediu, cum ar fi pielea, gura, cavitatea, esofagul și plămânul . Într-o etapă anterioară, SCC este însoțită de keratinizarea epidermică și formarea ulcerului în suprafața mucoasei, iar țesuturile mai profunde sunt invadate de SCC într-o etapă ulterioară. Pentru aspectul microscopic, celulele SCC dispuse în cuiburi, care sunt înconjurate de celule poligonale cu granițe celulare distincte și nuclee hipercromatice. CSSC au o tendință ridicată de a metastaza, de obicei prin ganglionii limfatici regionali, și ar putea provoca daune sistematice ale mai multor organe. Chirurgia, terapia cu laser și radiațiile continuă să fie cel mai utilizat tratament pentru CSSC . Caracteristicile citomorfologice unice ale variantelor SCC conduc la un tratament și rezultate distincte .
antigenul de suprafață celulară CD109 este o glicoproteină legată de glicozilfosfatidilinozitol (GPI) de aproximativ 170 kDa și un membru al familiei de proteine care conțin tioester a2 macroglobulină (a2m) / C3, C4, C5 . Gena CD109 umană este localizată în cromozomul 6Q, constituind aproximativ 3,3% din secvența genomică totală CD109 cu cei 33 de exoni ai săi . Cd109 ADNc izolat cuprinde un cadru de citire deschisă de 4335 bp care codifică un aminoacid 1445 (aa) . Proteina CD109 conține o peptidă lider N-terminală 21 aa, o regiune momeală presupusă (aa 651-683), un situs de legare tioester (aa 918-924), o hexapeptidă care definește reactivitatea tioester (aa1030–1035) și o secvență de semnal GPI-ancoră consens C-terminal cu clivajul prevăzut să apară după aminoacidul 1420 (Fig. 1) . Peptida lider N-terminală ancorează proteina la membrana interioară din periplasmă și fragmentul n-terminal al CD109 secretat din celule după scindare de către furin protează . Clivajul proteazei unei regiuni de momeală în mare parte dezordonate activează CD109, rezultând o schimbare conformațională care prinde proteaza într-o structură asemănătoare cu cușca și expune legătura tioester foarte reactivă. Situsul de legare tioester care, la activare, poate lega covalent CD109 de reziduurile de lizină de pe suprafața proteazei atacante. În plus, secvența semnalului GPI-ancoră consens C-terminal este, de asemenea, cunoscută sub numele de domeniu de legare a receptorilor .

The structure of CD109 protein. Lungimea completă a proteinei CD109 cuprinde 1445 aminoacizi (aa), inclusiv o peptidă de semnal n-terminal 21 aa, o regiune de momeală presupusă (aa 651-683), un situs de legare a tioesterului (aa 918-924), o hexapeptidă care definește reactivitatea tioester (aa1030–1035) și o secvență de semnal GPI-ancoră de consens C-terminal (aa 1420-1445) cu clivajul prevăzut 1420
cd109 a fost identificat pentru prima dată ca antigen de suprafață celulară printr-un anticorp monoclonal crescut împotriva liniei celulare limfoide/mieloide primitive kg1a . Studiile anterioare au raportat că CD109 este exprimat pe un subset de celule mononucleare ale măduvei osoase fetale și adulte CD34+ pozitive, limfoblaste t activate, trombocite activate, celule endoteliale , subseturi de celule stem mezenchimale și mai multe linii celulare tumorale umane, dar care nu sunt exprimate în celule T umane în repaus, trombocite sau leucocite din sângele periferic . În special, CD109 a fost detectat în SCC-urile mai multor organe, inclusiv plămânul , esofagul , colul uterin , pielea , penisul , cavitatea bucală și vezica biliară . CD109 este o componentă a sistemului receptor al factorului de creștere transformant-ctf1 (TGF-ctf1) și reglează negativ semnalizarea TGF-ctf1 . Pierderea și câștigul funcției studiile au sugerat că CD109 poate fi o țintă moleculară pentru dezvoltarea de noi terapii pentru CSSC de diferite origini tisulare . În această revizuire, vom prezenta un rezumat al cunoștințelor actuale despre CD109 și relația sa cu CSSC.
CD109 expresia în CSSC
în țesuturile tumorale, CD109 a fost detectat imunohistochimic în CSSC , precum și carcinoame uroteliale , melanoame maligne , carcinoame mamare bazale , mixofibrosarcom, sarcoame epiteliale și gliom . În special, Shiraki și colab. raportate celule tumorale perivasculare CD109-pozitive în țesuturile gliomului uman de grad inferior și într-un model de șoarece recapitulat gliom uman, sugerând un rol cheie al CD109 pentru această boală. Studiile anterioare au arătat că expresia ridicată a CD109 în CSSC și expresia limitată în celulele scuamoase normale (Tabelul 1) . Mai mult, CD109 este foarte exprimat în CSSC bine diferențiate, mai degrabă decât în CSSC moderat sau slab diferențiate, astfel nivelul de Expresie al CD109 este invers corelat cu gradul tumorii .
CD109 și SCC pulmonar
carcinomul cu celule scuamoase pulmonare (lscc) este unul dintre subtipurile majore ale-cancer pulmonar celular (NSCLC) și reprezintă aproximativ 20-30% din cazurile de NSCLC . Expresia CD109 a fost examinată în carcinoamele cu celule pulmonare umane prin RT-PCR cantitativ, care a arătat o expresie semnificativ mai mare a CD109 în carcinoamele cu celule scuamoase, dar nu și în adenocarcinoame, carcinoame cu celule mari sau carcinoame cu celule mici . În plus, Sato și colab. a folosit un anticorp CD109 pentru a analiza expresia CD109 în țesuturile normale de carcinom pulmonar și pulmonar. În țesuturile pulmonare normale, expresia CD109 a fost limitată în principal la celulele bazale ale epiteliilor bronșice și bronhiolare . În țesuturile carcinomului pulmonar, imunoreactivitatea puternică a fost detectată preferențial în Lscc, dar nu și în alte tipuri de carcinoame pulmonare . Aceste constatări indică faptul că CD109 este reglementat în mod specific în LSCCs.
CD109 și SCC esofagian
carcinomul cu celule scuamoase esofagiene (ESCC) reprezintă o șesime din totalul mortalității legate de cancer, care apare la o incidență mai mare în țările asiatice . Prognosticul ESCC rămâne slab, iar rata de supraviețuire de 5 ani a ESCC este de aproximativ 14-22% . Studii recente sugerează că expresia alterată a receptorilor TGF-XV contribuie la progresia ESCC, iar expresia crescută a proteinelor inhibitoare ale semnalizării TGF-XV se corelează cu prognosticul slab al ESCC . Hashimoto și colab. expresia CD109 examinată în țesuturile ESCC prin RT-PCR cantitativ, iar transcrierea ARNm CD109 a fost reglată în sus în aproximativ 50% din Escc. Ulterior, nivelul proteic al CD109 în ESCC a fost examinat prin imunohistochimie pe microarrays tisulare (TMA) . În țesutul esofagian non-bolnav, expresia CD109 a fost restricționată în citosolul celulelor epiteliale stratificate la un nivel slab. În toate probele ESCC din TMA, firele de celule epiteliale scuamoase maligne au prezentat o colorare puternică CD109 în citosol. Nu s-a observat nici o colorare CD109 pozitivă la alte tipuri de celule. Mai mult, expresia CD109 a fost mai mare la Escc – urile bine și moderat diferențiate decât la cele slab diferențiate .
CD109 și col uterin SCC
de col uterin (col uterin) cancer de conturi pentru aproximativ 12% din decese legate de cancer de femei . CSSC reprezintă 80-85% din toate cazurile de cancer de col uterin . Zhang și colab. a arătat că expresia CD109 a fost semnificativ mai mare în CSSC de col uterin decât în adenocarcinoamele endometriale. De asemenea, au investigat expresia CD109 în cinci linii celulare de carcinom cervical uman și au observat niveluri ridicate de Expresie CD109 în două linii celulare SCC.
CD109 și SCC cutanat
carcinomul cutanat cu celule scuamoase (CSCC) este al doilea tip cel mai frecvent de cancer de piele non-melanom (NMSC) cu o incidență în continuă creștere . Deși CSCC au un prognostic general favorabil, există încă aproximativ 1,5–2% dintre pacienți mor din cauza acestei boli . Expresia CD109 în CSCS a fost examinată prin imunohistochimie pe TMA . În piei normale, CD109 a fost slab exprimat în stratul bazal al celulelor epidermice, în timp ce firele de celule epidermice scuamoase maligne au prezentat o colorare puternică CD109. În plus, expresia CD109 a fost invers corelată cu gradele CSCC. La fel ca Escc – urile, expresia CD109 a fost mai mare la CSC-urile bine și moderat diferențiate decât la cele slab diferențiate . Carcinomul cu celule scuamoase peniene (PSCC) este un subtip de CSCC și a fost rar studiat . Dong și colab. a demonstrat că proteina CD109 este foarte exprimată în celulele scuamoase maligne ale PSCCs comparativ cu țesuturile normale ale penisului pe TMA. Mai mult, modelul de Expresie este validat pe probe PSCC chirurgicale proaspete prin imunofluorescență, qRT-PCR și Western blotting, sugerând că CD109 poate fi un biomarker pentru PSCC .
CD109 și cavitatea orală SCC
carcinomul cu celule scuamoase orale (OSCC) reprezintă 2-3% din toate cazurile de cancer la nivel mondial . Pierderea mutației funcției în receptorul de tip II TGF-XV este un eveniment frecvent pentru cavitatea bucală SCC . Expresia CD109 în țesuturile orale normale și țesuturile OSCC de la 124 de pacienți a fost examinată prin colorare imunohistochimică. Niveluri ridicate de Expresie CD109 au fost detectate frecvent în CSSC și leziunile premaligne ale cavității bucale, dar nu și în epiteliile scuamoase normale . Mai mult, nivelul de Expresie al CD109 a fost semnificativ mai mare în OSCCs bine diferențiate decât în OSCCs moderat sau slab diferențiate, ceea ce implică faptul că expresia CD109 este corelată cu etapele de diferențiere ale OSCCs . În plus , liniile celulare OSCC supraexprimând CD109 au prezentat o creștere accelerată a celulelor in vitro, implicând faptul că CD109 implică progresia OSCCs.
CD109 și SCC vezicii biliare
Cancerul vezicii biliare (GBC) este cel mai agresiv dintre cancerele biliare cu cea mai scurtă supraviețuire mediană . Carcinomul cu celule scuamoase ale vezicii biliare (GSCC) reprezintă 3% din neoplasmul malign al acestui organ . Subtipurile de țesuturi GBCs, inclusiv adenocarcinomul (AC), carcinomul cu celule scuamoase (SCC) și carcinomul adenosquamos (ASC) au fost examinate pe TMA prin colorare imunohistochimică cu un anticorp CD109. Colorația CD109 a fost negativă în toate țesuturile normale ale vezicii biliare și în țesuturile AC. Între timp, celulele CD109 pozitive au fost găsite în 86,7% din SCC și 91,7% din ASCs. Deoarece CD109 este exprimat distinct în celulele scuamoase maligne din vezica biliară, CD109 poate fi un marker de diagnostic pentru SCC-urile vezicii biliare și ASCs .
calea de semnalizare TGF-XV și SCCs
calea de semnalizare TGF-XV este implicată în multe procese celulare, inclusiv creșterea celulară, apoptoza diferențierii celulare și homeostazia celulară . Familia TGF-β liganzi, TGF-β1, TGF-β2 și TGF-β3, se leagă specific transmembranar de tip I și de tip II serin/treonin kinaza receptori (TGF-ßR1 și TGF-ßR2) , rezultând în activarea TGF-ßR1 activitatea kinazei . TGF-ecacr1 activat propagă apoi semnalul prin fosforilarea substraturilor sale intracelulare, R-SMADs (Smad2 și Smad3) . Smad2 și Smad3 interacționează cu TGF-ectr1 și SARA (ancoră Smad pentru activarea receptorului), o proteină din domeniul FYVE care interacționează direct cu Smad2 și Smad3, SARA funcționează pentru a recruta Smad2 la receptorul TGF-XV , apoi r-Smad fosforilat formează complexe heteromerice cu Co-SMAD (Smad4) . După fosforilarea și complexele ulterioare cu Smad4, aceste complexe R-Smads sunt eliberate din TGF-inkrtr1 și SARA , apoi se translocează în nucleu unde interacționează cu factorii de transcripție care îi recrutează la elemente promotoare specifice ale genelor țintă .
Endocitoza receptorilor este un mecanism de reglare pivot în transducția semnalului. Receptorii TGF-XV sunt interiorizați atât prin căi dependente de clatrin, cât și de caveolae. Interiorizarea receptorilor TGF-XV prin gropile acoperite cu clatrin a fost legată de semnalizarea prin Smad2/3 și reciclarea receptorilor. În schimb, localizarea receptorului TGF-XV în caveolae este asociată cu reglarea descendentă a semnalizării Smad2/3 și degradarea receptorului în urma ubiquitinării de către E3-ubiquitin ligază Smurf2 . Cu toate acestea, Smad-urile inhibitoare (Smad6 și Smad7) formează o subclasă distinctă de Smad-uri care acționează într-o manieră opusă R-Smad-urilor și antagonizează semnalizarea . Acestea pot concura cu R-Smad – uri pentru legarea la TGF-ectrocr1 activat și, astfel, pentru a inhiba fosforilarea R-Smad-urilor . În plus, recrutează ligaze E3-ubiquitin la TGF-inkrtr1 activat, rezultând ubiquitinarea și degradarea receptorilor și încetarea semnalizării .
dereglarea căii TGF-XV a fost implicată în mai multe tipuri de cancer . Studiile au demonstrat că semnalizarea TGF-XV provoacă un efect preventiv în stadiile anterioare ale tumorigenezei, dar un efect supresiv în timpul dezvoltării ulterioare a tumorii . Mutațiile genei TGF-ectr1 au fost, de asemenea, găsite în SCC-urile pielii, sugerând că inactivarea TGF-XV duce la inițierea SCC-urilor .
CD109 și calea de semnalizare TGF-Coloseni
CD109 este un co-receptor TGF-colosali și modulează activitatea receptorului de semnalizare TGF-centuri într-o manieră specifică celulei . Pe suprafața celulară, CD109 reglează negativ calea de semnalizare TGF-XV1 prin formarea unui complex receptor cu TGF-ecacr1 și TGF-eccr2 în keratinocitele umane . Receptorii TGF-XV se degradează după ubiquitinare de către E3-ubiquitin ligază Smurf2, și sunt interiorizați atât prin căi dependente de clatrin, cât și prin căi dependente de caveolae . Bizet și colab. s-a demonstrat că CD109 se asociază cu caveolin-1 și promovează endocitoza receptorului TGF-XV. În plus, CD109 promovează localizarea receptorilor TGF-XV în compartimentul caveolar în prezența ligandului și facilitează degradarea receptorului TGF-XV. CD109 reglementează, de asemenea, localizarea și asocierea Smad7/Smurf2 cu TGF-ecacr1. Efectele inhibitoare ale CD109 necesită expresia Smad7 și activitatea smurf2 ubiquitin ligase . Mai mult, CD109 poate fi eliberat de pe suprafața celulară prin lipaze celulare, cum ar fi fosfatidilinozitol-fosfolipaza c specifică (PI-PLC). Forma solubilă a CD109 își păstrează capacitatea de a se lega TGF-XV1 și de a-l confisca departe de receptorii TGF-XV .
cu toate acestea, Vorstenbosch și colab. a raportat că CD109 reglementate diferențiat induse de TGF-XV ALK1-Smad1 / 5 față de căile ALK5-Smad2/3 (ALK1 și ALK5 sunt toți receptori de tip I TGF-XV). Ei au descoperit că semnalizarea TGF-IX inhibă proliferarea și migrarea celulelor endoteliale, în timp ce semnalizarea TGF-x x x induce, de asemenea, aceste procese prin ALK1-Smad1/5 . Ei demonstrează că ALK1 este exprimat și co-localizează cu CD109 în keratinocitele de șoarece și că șoarecii care supraexprimă CD109 în epidermă prezintă o semnalizare îmbunătățită ALK1-Smad1/5, dar au scăzut semnalizarea ALK5-Smad2/3 .
În plus, TGF-XV1 este un inhibitor puternic al creșterii în majoritatea celulelor epiteliale . Hagiwara și colab. s-a demonstrat că supraexprimarea liniilor celulare SCC pe cale orală CD109 a accelerat proliferarea celulară și a afectat efectul antiproliferativ mediat de TGF-XV1. În schimb, celulele SCC cu knockdown CD109 au prezentat o creștere mai lentă a celulelor . Un nivel ridicat al expresiei CD109 a inhibat fosforilarea Smad2, atenuând astfel semnalizarea TGF-ctf1 / Smad2 și afectează suprimarea mediată de TGF-ctf1 a creșterii celulare, CD109 knockdown a crescut fosforilarea Smad2 prin stimularea TGF-ctf1 . Deși CD109 reglementează și semnalizarea Smad1 / 5, nu a fost legată de dezvoltarea SCC. Împreună, CD109 facilitează dezvoltarea SCC-urilor prin inhibarea căii TGF-XV-Smad2 / 3 (Fig. 2).

cd109 și TGF-semnalizare la nivel mondial. TGF-XV leagă TGF-oktogr2, care recrutează și fosforilează TGF-oktogr1. Activat TGF-ecacr1 propagă semnalul în aval prin fosforilarea directă a Smad2 și Smad3. R-Smad-urile fosforilate formează apoi complexe heteromerice cu Smad4, combinate cu factori de transcripție, reglează transcripția genei și funcția celulară. Receptorii TGF-XV se interiorizează prin gropile acoperite cu clatrin sau prin calea caveolară. CD109 crește legarea TGF-VIII la receptorii TGF-VIII și promovează localizarea receptorului TGF-VIII la caveolae, în cele din urmă crește endocitoza receptorului TGF-VIII și facilitează degradarea receptorului TGF-VIII. Efectele inhibitoare ale CD109 necesită expresia Smad7 și activitatea smurf2 ubiquitin ligase
CD109 și semnalizarea STAT3
traductorul de semnal și activatorul factorului de transcripție 3 (STAT3) este esențial pentru proliferarea, diferențierea, migrarea, supraviețuirea și oncogeneza celulelor . Litvinov și colab. a raportat că expresia proteinei CD109 a scăzut semnificativ în epiderma psoriazică în comparație cu pielea adiacentă neimplicată. Cu toate acestea, expresia ARNm CD109 este neschimbată în plăcile psoriazice în comparație cu pielea normală, sugerând posibilitatea ca eliberarea de proteine CD109 să fie îmbunătățită în keratinocitele psoriazice . Ei au sugerat că cd109 eliberat / solubil este capabil să inducă modificări moleculare despre care se știe că apar în psoriazis . In vitro, au descoperit că transfecția CD109 siRNA reglează în jos STAT3, eliberarea CD109 de pe suprafața celulară a keratinocitelor umane cultivate. În plus, CD109 exogen/recombinant induce semnalizarea STAT3 în keratinocitele umane . În plus, Chuang și colab. a raportat că expresia CD109 a fost reglată dramatic în celulele adenocarcinomului pulmonar metastatic, iar celulele care exprimă un cd109 shRNA (shCD109) au arătat o reducere dramatică a fosforilării STAT3. STAT3 knockdown a redus foarte mult metastazele, iar restaurarea activității STAT3 a crescut capacitatea celulelor care exprimă shCD109 de a metastaza . La activare, STAT3 este fosforilat de proteina non-receptor tirozin kinaze janus kinaza 2 (JAK2), ducând la formarea dimerului STAT3 și translocarea în nucleu . Cu toate acestea, inhibarea activității jak kinazei în fibroblastele care supraexprimă CD109 a redus starea fosforilată3 la un nivel similar cu cel din celulele parentale care exprimă niveluri scăzute de CD109, sugerând că fosforilarea STAT3 indusă de CD109 necesită activitate jak kinazei. Astfel, semnalizarea JAK / STAT3 ar putea media efectele CD109 în creșterea tumorii și metastaze .
deși eliminarea CD109 în keratinocitele umane și celulele adenocarcinomului pulmonar reglează în jos semnalizarea STAT3 in vitro , șoarecii cu deficit de CD109 au prezentat rezultate opuse. Mii și colab. au generat șoareci cu deficit de CD109, care au prezentat anomalii ale pielii, inclusiv hiperplazie epitelială și infiltrare celulară inflamatorie. Ei au raportat că fosforilarea STAT3 la șoarecii cu deficit de CD109 a fost semnificativ mai mare comparativ cu șoarecii de tip sălbatic. În plus, reglarea în sus a semnalizării STAT3 este asociată cu proliferarea crescută și diferențierea afectată a keratinocitelor .
discrepanța rezultatelor din studiile in vitro și in vivo ar putea fi cauzată de modificările sistematice ale micromediului în țesuturile șoarecilor cu deficit de CD109. Pierderea CD109 în toate celulele la șoareci ar putea modifica micromediul subcutanat care activează semnalizarea STAT3 în keratinocite. În plus, CD109 poate exercita efecte de reglementare distincte în diferite tipuri de celule, ducând la modificări specifice tipului de celulă în semnalizarea STAT3. În plus față de keratinocite, CD109 este exprimat în celule endoteliale, celule epiteliale și fibroblaste, care participă la constituie țesutul pielii. Cu toate acestea, până în prezent relația dintre semnalizarea CD109 și STAT3 nu a fost explorată în aceste tipuri de celule.
CD109 și semnalizarea EGFR
receptorul factorului de creștere Epidermal (EGFR) este un membru al familiei de receptori ErbB. La legarea ligandului prin EGF, EGFR formează dimeri, fie homodimeri, fie heterodimeri cu un alt membru al familiei ErbB HER2 . Receptorii dimerizați se auto-fosforilează reciproc și apoi fosforilează proteina non-receptor tirozină c-src kinază, care activează STAT3 . Activarea EGFR promovează migrarea, supraviețuirea și proliferarea celulelor. In tumorile maligne EGFR supra-exprimare este corelată cu adâncimea de invazie a tumorii și legate de prognostic mai saraci . Mutațiile care duc la supraexprimarea EGFR sunt detectate în SCC pulmonar , SCC cap și gât și SCC esofag . CD109 ancorat cu membrană în celulele SK-MG-1 interacționează direct cu EGFR și îmbunătățește semnalizarea EGF, ceea ce crește ulterior migrarea și invazia celulelor, în timp ce cd109 secretat nu are niciun efect asupra semnalizării EGF . EGFR ar putea media efectele CD109 asupra semnalizării STAT3, care necesită studii suplimentare pentru a elucida (Fig. 3).
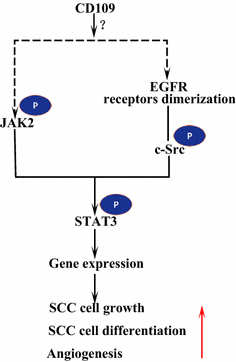
CD109 și EGFR / STAT3 semnalizare. CD109 facilitează semnalizarea EGFR. Receptorii EGFR dimerizați fosforilează c-Src, care la rândul său fosforilează STAT3. CD109 poate, de asemenea, activează STAT3 prin fosforilarea JAK2, apoi promovează creșterea celulelor SCC, diferențierea și angiogeneza
până în prezent imaginea de ansamblu a tumorigenezei și dezvoltarea SCC rămân incomplete. Există discuții încrucișate între căile de semnalizare TGF-XV și STAT3 sau EGFR. CD109 este probabil unul dintre efectorii cheie ai rețelei de semnalizare care reglează SCC-urile. Cu toate acestea, nu s-au raportat dovezi directe care să delimiteze rolurile CD109 în patogeneza CSSC.
perspective
studiile din probe de țesut uman indică faptul că CD109 este foarte exprimat în CSSC ale mai multor organe, în special în celulele scuamoase maligne bine diferențiate . Prin Detectarea expresiei CD109 cu imunohistochimie în țesuturile umane, CD109 poate acționa potențial un biomarker pentru a determina progresia SCC. Studiile actuale sugerează că CD109 este foarte exprimat în CSC bine diferențiate, iar expresia sa este mai mică în CSC nediferențiate . Cu toate acestea, nu este clar dacă CD109 este asociat cu invazia vasculară, metastazele și prognosticul după operație. Prin urmare, sunt necesare studii suplimentare pentru a explora semnificația clinicopatologică a CD109 în CSSC într-o dimensiune mai mare a eșantionului.CD109 este o glicoproteină legată de GPI, care îi permite să fie eliberată din membrană . Forma solubilă a CD109 afectează, de asemenea, legarea TGF-XV la receptorii săi și, ulterior, modulează progresia SCC . Litvinov și colab. s-a constatat că CD109 eliberat de pe suprafața celulei în mediul extracelular, iar forma eliberată de CD109 își păstrează capacitatea de a induce căi de semnalizare intracelulară. În plus, Sakakura și colab. a raportat că CD109 ser a fost eliberat de tumora xenogrefă și crește proporțional cu volumul de xenogrefă tumorii. Prin urmare, detectarea nivelului seric CD109 ar putea ajuta la monitorizarea tumorilor care supraexprimă CD109, inclusiv CSSC.
Exozomii comunică leziunea tumorală primară și nișa sa prin pachetul său care conține proteine selectate sau alte molecule . Izolarea și analizele exozomului asociat tumorii circulante pot servi drept biomarkeri pentru diagnosticarea pacienților cu cancer. CD109 exogen a fost identificat ca o componentă a exozomului secretat din 293 de celule transfectate, făcându-l o țintă promițătoare pentru diagnosticul exozomului . Totuși, până în prezent CD109 nu au fost raportate în exozomi derivați din SCC sau alte celule tumorale. Mecanismele care stau la baza ambalării CD109 în exozomi merită o investigație mai detaliată.
în plus față de potențialul ca biomarker, CD109 ar putea fi o țintă pentru abordările terapeutice. CD109 este o proteină membranară, care poate fi vizată direct de anticorpi sau enzime specifice. CD109 ar putea fi, de asemenea, recunoscut de vehiculele vizate pentru livrarea de droguri. Cu toate acestea, rolurile detaliate ale CD109 în patogeneza CSSC sunt încă neclare. De exemplu, trebuie definită relația dintre expresia inferioară a CD109 și CSSC nediferențiate. Intervenția CD109 poate fi luată în considerare în practica clinică numai după ce riscurile și beneficiile sunt evaluate cu atenție.