
carcinoma de células Escamosas (SCC), também conhecido como espinho carcinoma de células, é uma das principais causas de câncer relacionados com morte em todo o mundo. SCC é um tumor epitelial maligno, surgindo em tecidos que fornecem uma barreira entre um organismo e o ambiente, como a pele, oral, cavidade, esôfago e pulmão . Em uma fase anterior, a CCSC é acompanhada por queratinização epidérmica e formação de úlceras na superfície da mucosa, e os tecidos mais profundos são invadidos pelo CCSC em uma fase posterior. Para aparência microscópica, as células SCC dispostas em ninhos, que são cercadas de células poligonais com fronteiras celulares distintas e núcleos hipercromáticos. Os SCCs têm uma alta tendência para metástases, geralmente através de gânglios linfáticos regionais, e pode causar danos sistemáticos de múltiplos órgãos. Cirurgia, terapia laser e radiação continuam a ser o tratamento mais amplamente utilizado para SCCs . As características citomorfológicas únicas das variantes SCC levam a tratamentos e resultados distintos .
o antigénio de superfície celular CD109 é uma glicoproteína ligada a glicosilfosfatidilinositol (GPI) de aproximadamente 170 kDa e membro da família a2 macroglobulina (A2M)/C3, C4, C5 de proteínas contendo tioéster . O gene CD109 humano está localizado no cromossomo 6q, constituindo aproximadamente 3,3% da sequência genômica CD109 total com seus 33 exons . O cd109 cDNA isolado inclui um quadro de leitura aberta de 4335 bp que codifica um aminoácido 1445 (aa). A proteína CD109 contém um peptídeo líder N-terminal de 21 aa, uma região de isca putativa (aa 651-683), um local de ligação do tioéster (aa 918-924), uma reatividade do tioéster definindo hexapéptido (aa1030–1035) e uma sequência de sinal de âncora GPI de consenso C-terminal com a clivagem prevista para ocorrer após o aminoácido 1420 (Fig. 1) . O péptido-líder N-terminal prende a proteína à membrana interna no periplasma e ao fragmento N-terminal do CD109 secretado das células após clivagem pela protease da furina . A clivagem da protease de uma região de isca em grande parte desordenada ativa o CD109, resultando em uma mudança conformacional que prende a protease em uma estrutura semelhante à gaiola e expõe a ligação de tioéster altamente reativa. O sítio de ligação do tioéster que, na activação, pode ligar covalentemente o CD109 aos resíduos de lisina na superfície da protease atacante. Além disso, a sequência de sinal de âncora GPI-consenso C-terminal também é conhecida como o domínio de ligação ao receptor .

The structure of CD109 protein. O comprimento total de CD109 proteína compreende 1445 de aminoácidos (aa), incluindo um 21 aa N-terminal do peptídeo sinal, um suposto isca região (aa 651-683), um thioester sítio de ligação (aa 918-924), um thioester reatividade definição hexapeptide (aa1030–1035) e um C-terminal consenso GPI-âncora da sequência de sinais (aa 1420-1445) com a clivagem prevista para ocorrer depois de aminoácidos 1420
CD109 foi identificado pela primeira vez como uma célula-superfície do antígeno por um anticorpo monoclonal levantadas contra a primitiva linfóides/mielóide linha celular KG1a . Estudos anteriores relataram que o CD109 é expresso num subconjunto de células mononucleares da medula óssea positiva do feto e do adulto CD34+, linfoblastos t activados, plaquetas activadas, células endoteliais, subconjuntos mesenquimais de células estaminais e várias linhas de células tumorais humanas , mas não expresso em células T humanas em repouso, plaquetas ou leucócitos do sangue periférico . Em particular, o CD109 foi detectado em SCCs de vários órgãos, incluindo pulmão , esófago , colo uterino , Pele , Pénis , cavidade oral e vesícula biliar . CD109 é um componente do sistema receptor do fator de crescimento transformador-β1 (TGF-β1) e regula negativamente a sinalização TGF-β1 . Perda e ganho de estudos de função sugeriu que o CD109 pode ser um alvo molecular para o desenvolvimento de novos terapeutas para SCCs de várias origens de tecidos . Nesta revisão, apresentaremos um resumo do conhecimento atual do CD109 e sua relação com o CCSC.
expressão CD109 no SCCs
nos tecidos tumorais, o CD109 foi imunohistocemicamente detectado no SCCs , bem como carcinomas uroteliais , melanomas malignos , carcinomas da mama tipo basal , mixofibrosarcoma, sarcomas epiteliais e glioma . Em particular, Shiraki et al. as células tumorais perivasculares CD109 notificadas em tecidos de glioma de grau inferior humano e num modelo de ratinho recapitularam o glioma humano, sugerindo um papel fundamental do CD109 para esta doença. Estudos anteriores demonstraram que a alta expressão do CD109 no SCCs e a limitada expressão em células escamosas normais (Tabela 1) . Além disso, o CD109 é altamente expresso em SCCs bem diferenciados e não em SCCs moderadamente ou mal diferenciados, assim o nível de expressão do CD109 é inversamente correlacionado com o grau tumoral .
CD109 e pulmão SCC
Pulmão, carcinoma de células escamosas (LSCC) é um dos principais subtipos de não-pequenas-câncer de pulmão de células (NSCLC), e representa cerca de 20% a 30% de casos de NSCLCs . A expressão CD109 foi examinada em carcinomas de células pulmonares humanas por RT-PCR quantitativa, que mostrou uma expressão significativamente mais elevada de CD109 em carcinomas de células escamosas, mas não em adenocarcinomas, carcinomas de grandes células ou carcinomas de pequenas células . Além disso, Sato et al. usou um anticorpo CD109 para analisar a expressão CD109 em tecidos normais de carcinoma pulmonar e pulmonar. Nos tecidos pulmonares normais, a expressão CD109 foi confinada principalmente às células basais do epitélio brônquico e bronquiolar . Nos tecidos do carcinoma do pulmão, a imunoreactividade forte foi detectada preferencialmente em LSCCs, mas não em outros tipos de carcinomas pulmonares . Estas constatações indicam que o CD109 está especificamente regulamentado em LSCCs.o carcinoma de células escamosas do esófago (ESCC) é responsável por um sexto de toda a mortalidade relacionada com o cancro, ocorrendo com uma incidência mais elevada nos países asiáticos . O prognóstico da ESCC permanece fraco, e a taxa de sobrevivência de 5 anos da ESCC é de cerca de 14-22% . Estudos recentes sugerem que a expressão alterada dos receptores TGF-β contribui para a progressão ESCC, e a expressão elevada das proteínas inibitórias da sinalização TGF-β correlaciona-se com o mau prognóstico da ESCC . Hashimoto et al. examined CD109 expression in ESCC tissues by quantitative RT-PCR, and CD109 mRNA transcription was upregulated in about 50% of the ESCCs. Mais tarde, o nível proteico do CD109 no ESCC foi examinado pela imunohistoquímica em microarrays tecidulares (TMA) . No tecido esofágico não doente, a expressão CD109 foi restringida no citosol das células epiteliais estratificadas a um nível fraco. Em todas as amostras ESCC dos TMAs, as cadeias de células epiteliais escamosas malignas apresentavam uma forte coloração CD109 no citosol. Não foi observada coloração CD109 positiva noutros tipos de células. Além disso, a expressão CD109 foi mais elevada em Escc bem diferenciadas e moderadamente diferenciadas do que nos Escc mal diferenciados .o cancro do colo do útero (colo do útero) é responsável por cerca de 12% da morte das mulheres relacionada com o cancro. Os SCC são responsáveis por 80-85% de todos os cancros cervicais . Zhang et al. demonstrou que a expressão do CD109 foi significativamente mais elevada nos SCCs cervicais do que nos adenocarcinomas do endométrio. Eles também investigaram a expressão do CD109 em cinco linhas celulares de carcinoma do colo do útero humano, e observaram níveis elevados de expressão do CD109 em duas linhas celulares SCC.
CD109 e CSC cutânea
carcinoma cutâneo de células escamosas (CSCC) é o segundo tipo mais comum de cancro de pele não melanoma (NMSC) com uma incidência constantemente crescente . Embora a CSCC tenha um prognóstico geralmente favorável, ainda há aproximadamente 1,5-2% dos pacientes morrem desta doença . A expressão CD109 em CSCS foi examinada por imunohistoquímica em TMAs . Nas peles normais, o CD109 foi fracamente expresso na camada basal de células epidérmicas, enquanto as cadeias de células epidermais malignas apresentavam uma forte coloração CD109. Além disso, a expressão CD109 estava inversamente correlacionada com as notas CSCC. Tal como o Escc, a expressão do CD109 foi mais elevada nos CSCC bem diferenciados e moderadamente diferenciados do que nos CSC pouco diferenciados . O carcinoma de células escamosas do pénis (PSCC) é um subtipo de CSCCs e raramente foi estudado . Dong et al. demonstrou que a proteína CD109 é altamente expressa em células escamosas malignas de PCSC em comparação com os tecidos penianos normais em TMAs. Além disso, o padrão de expressão é validado em amostras de PSCC cirúrgicas frescas por imunofluorescência, qRT-PCR e western blotting, sugerindo que CD109 pode ser um biomarcador para PSCC .o carcinoma de células escamosas orais (OSCC) é responsável por 2-3% de todos os cancros em todo o mundo . A perda de mutação da função no receptor tipo II TGF-β é um acontecimento frequente na cavidade oral SCC . A expressão CD109 nos tecidos orais normais e nos tecidos OSCC de 124 doentes foi examinada por coloração imunohistoquímica. Foram frequentemente detectados níveis elevados de expressão CD109 em CCSC e lesões pré-malignas da cavidade oral, mas não em epitélios escamosos normais . Além disso, o nível de expressão do CD109 foi significativamente mais elevado nos OSCCs bem diferenciados do que nos OSCCs moderadamente ou mal diferenciados, o que implica que a expressão do CD109 está correlacionada com as fases de diferenciação dos OSCCs . Além disso , as linhas celulares OSCC sobreexpressing CD109 exibiram crescimento celular acelerado in vitro, implicando que CD109 envolve na progressão dos OSCCs.o cancro da vesícula biliar (GBC) é o mais agressivo dos cancros biliares com menor sobrevida mediana . O carcinoma de células escamosas da vesícula biliar (GSCC) é responsável por 3% do neoplasma Maligno deste órgão . Os subtipos de tecidos GBCs, incluindo adenocarcinoma (AC), carcinoma de células escamosas (SCC) e carcinoma adenosquâmico (ASC), foram examinados em TMAs por coloração imunohistoquímica com um anticorpo CD109. A coloração CD109 foi negativa em todos os tecidos normais da vesícula biliar e tecidos AC. Enquanto isso, células CD109 positivas foram encontradas em 86,7% do SCCs e 91,7% do ASCs. Como o CD109 é claramente expresso em células sqamosas malignas na vesícula biliar, o CD109 pode ser um marcador de diagnóstico para SCCs e ASCs da vesícula biliar .
via de sinalização TGF-β e SCCs
a via de sinalização TGF-β está envolvida em muitos processos celulares, incluindo crescimento celular, apoptose de diferenciação celular e homeostase celular . A família de ligantes TGF-β, TGF-β1, TGF-β2 e TGF-β3 liga-se a receptores transmembranares específicos do tipo I e da serina/cinase da treonina do tipo II (TGF-ßR1 e TGF-ßR2) , resultando na activação da actividade cinase TGF-ßR1 . O TGF-ßR1 activado propaga então o sinal através da fosforilação dos seus substratos intracelulares, R-SMADs (Smad2 e Smad3) . Smad2 e Smad3 interagir com TGF-ßR1 e SARA (Smad âncora para a ativação do receptor), um FYVE domínio proteína que interage diretamente com Smad2 e Smad3, SARA funções para recrutar Smad2 para o TGF-β do receptor , em seguida, o fosforilada R-SMADs formulário heteromeric complexos com Co-SMAD (Smad4) . Após a fosforilação e complexos subsequentes com Smad4, estes complexos R-Smads são libertados de TGF-ßR1 e SARA, em seguida translocados para o núcleo onde interagem com factores de transcrição que os recrutam para elementos promotores específicos dos genes-alvo .a endocitose do Receptor é um mecanismo regulador central na transdução do sinal. Os receptores TGF-β são internalizados através das vias dependentes da clatrina e das caveolae. A internalização dos receptores TGF-β através das fossas revestidas de clatrina tem sido associada à sinalização via Smad2/3 e à reciclagem de receptores. Em contraste, a localização do receptor TGF-β nas caveolae está associada à redução da sinalização Smad2/3 e à degradação do receptor após ubiquitinação pela ligase E3-ubiquitina Smurf2 . No entanto, Smads inibitórios (Smad6 e Smad7) formam uma subclasse distinta de Smads que agem de uma maneira oposta a R-Smads e antagonizam a sinalização . Podem competir com R-Smads pela ligação ao TGF-ßR1 activado, inibindo assim a fosforilação do R-Smads . Além disso, recrutam ligases E3-ubiquitina para o TGF-ßR1 ativado, resultando em ubiquitinação e degradação do receptor, e fim da sinalização .a desregulação da via TGF-β tem sido implicada em vários tipos de cancro . Estudos têm demonstrado que a sinalização TGF-β provoca um efeito preventivo durante os estágios iniciais da tumorigénese, mas um efeito supressivo durante o desenvolvimento posterior do tumor . Foram também encontradas mutações no gene TGF-ßR1 no SCCs da pele, sugerindo que a inactivação do TGF-β conduz ao início do SCCs .
CD109 e a via de sinalização TGF-β
CD109 é um co-receptor TGF-β, e modula a actividade do receptor de sinalização TGF-β de uma forma específica da célula . Na superfície celular, o CD109 regula negativamente a via sinalizadora TGF-β1 através da formação de um complexo receptor com TGF-ßR1 e TGF-ßR2 em queratinócitos humanos . Os receptores TGF-β degradam-se após a ubiquitinação pela ligase E3-ubiquitina Smurf2 , e são internalizados através das vias dependentes da clatrina e das caveolae . Bizet et al. demonstrou que o CD109 associa-se à caveolin-1 e promove a endocitose do receptor TGF-B. Além disso, o CD109 promove a localização dos receptores TGF-β no compartimento caveolar na presença de ligando e facilita a degradação do receptor TGF-β. O CD109 também regula a localização e a associação do Smad7/Smurf2 com o TGF-ßR1. Os efeitos inibitórios do CD109 requerem expressão Smad7 e actividade da ligase ubiquitina do Smurf2 . Além disso, o CD109 pode ser libertado da superfície celular por lipases celulares, tais como fosfolipase c específica ao fosfatidilinositol (PI-PLC). A forma solúvel do CD109 mantém a sua capacidade de se ligar ao TGF-β1 e confiscá-lo dos receptores TGF-β .
no entanto, Vorstenbosch et al. reported that CD109 differentially regulated TGF-β-induced ALK1-Smad1/5 versus ALK5-Smad2/3 pathways (ALK1 and ALK5 are all TGF-β type I receptors). They found that TGF-β signaling inibe a proliferação e migração das células endoteliais, while TGF-β signaling also induces these processes via ALK1-Smad1 / 5 . Eles demonstram que ALK1 é expresso e co-localiza com CD109 no mouse queratinócitos e que os ratos overexpressing CD109 na epiderme de exibição avançado ALK1-Smad1/5 de sinalização, mas diminuiu ALK5-Smad2/3 de sinalização .além disso, TGF-β1 é um potente inibidor do crescimento na maioria das células epiteliais . Hagiwara et al. demonstrou que a sobreexpressão oral das linhas celulares SCC CD109 acelerou a proliferação celular e prejudicou o efeito anti-proliferativo mediado pelo TGF-β1. Em contraste, as células SCC com knockdown CD109 exibiram um crescimento celular mais lento . Um elevado nível de expressão do CD109 inibiu a fosforilação do Smad2, atenuando assim a sinalização TGF-β1 / Smad2 e prejudicando a supressão do crescimento celular mediada pelo TGF-β1, a neutralização do CD109 aumentou a fosforilação do Smad2 pela estimulação TGF-β1 . Embora o CD109 também regule a sinalização Smad1/5, ele não tem sido conectado com o desenvolvimento do SCC. Em conjunto, o CD109 facilita o desenvolvimento de SCCs através da inibição da via TGF-β-Smad2/3 (Fig. 2).

CD109 and TGF-β signaling. O TGF-β liga o TGF-ßR2, que recruta e fosforila o TGF-ßR1. O TGF-ßR1 activado propaga o sinal a jusante através da fosforilação directa do Smad2 e do Smad3. R-SMADs fosforilados formam então complexos heteroméricos com Smad4, combinados com fatores de transcrição, regulam a transcrição genética e a função celular. Os receptores TGF-β interiorizam-se através das fossas revestidas de clatrina ou da via caveolar. O CD109 aumenta a ligação do TGF-β aos receptores TGF-β e promove a localização do receptor TGF-β Para As caveolae, em última análise aumenta a endocitose do receptor TGF-β e facilita a degradação do receptor TGF-B. Os efeitos inibitórios da CD109 exigir expressão de Smad7 e Smurf2 hidrolase ligase atividade
CD109 e STAT3 sinalização
o Sinal do transdutor e ativador do fator de transcrição 3 (STAT3) é fundamental para a proliferação celular, diferenciação, migração, sobrevivência, e a oncogênese . Litvinov et al. relataram que a expressão da proteína CD109 foi significativamente diminuída na epiderme psoriática em comparação com a pele adjacente não envolvida. No entanto, a expressão de mRNA CD109 permanece inalterada em placas psoriáticas em comparação com a pele normal, sugerindo a possibilidade de que a libertação de proteína CD109 é aumentada em queratinócitos psoriáticos . Eles sugeriram que o CD109 liberado/solúvel é capaz de induzir alterações moleculares que são conhecidas por ocorrer na psoríase . In vitro, descobriram que a transfecção de siRNA CD109 regula o STAT3, libertação de CD109 da superfície celular de queratinócitos humanos cultivados. Além disso, o CD109 exógeno / recombinante induz a sinalização STAT3 nos queratinócitos humanos . Além disso, Chuang et al. informou que CD109 expressão foi dramático como potenciadas na metástase de adenocarcinoma de pulmão de células e células que expressam uma CD109 shRNA (shCD109) mostrou uma redução dramática na STAT3 fosforilação. STAT3 knockdown reduziu significativamente as metástases, e a restauração da atividade do STAT3 aumentou a capacidade das células que expressam shCD109 para metastizar . Após a ativação, o STAT3 é fosforilado pela proteína não-receptora tirosina cinases janus kinase 2 (JAK2), levando à formação do STAT3 dímero e translocação para o núcleo . No entanto, a inibição da actividade da JAK kinase nos fibroblastos que sobreexpressam o CD109 reduziu o STAT3 fosforilado para um nível semelhante ao das células parentais que expressam níveis baixos de CD109, sugerindo que a fosforilação do STAT3 induzida pelo CD109 requer a actividade da JAK kinase. Assim, a sinalização JAK/STAT3 pode mediar os efeitos do CD109 no crescimento tumoral e metástase .embora a diminuição do CD109 nos queratinócitos humanos e nas células do adenocarcinoma pulmonar regule in vitro o STAT3 sinalizando in vitro , os ratos com deficiência em CD109 apresentaram resultados opostos. Mii et al. ratos com deficiência em CD109, que exibiram anomalias cutâneas incluindo hiperplasia epitelial e infiltração celular inflamatória. Relataram que a fosforilação do STAT3 em ratinhos com deficiência em CD109 foi significativamente mais elevada em comparação com os ratinhos do tipo selvagem. Além disso, a regulação da sinalização do STAT3 está associada ao aumento da proliferação e à diferenciação deficiente dos queratinócitos .a discrepância dos resultados dos estudos in vitro e in vivo pode ser causada pelas alterações sistemáticas do microambiente nos tecidos de ratinhos com deficiência de CD109. A perda de CD109 em todas as células dos ratinhos pode modificar o microambiente subcutâneo que activa a sinalização do STAT3 nos queratinócitos. Além disso, o CD109 pode exercer efeitos regulatórios distintos em diferentes tipos de células, levando à modificação específica do tipo de célula na sinalização do STAT3. Para além dos queratinócitos, o CD109 é expresso em células endoteliais, células epiteliais e fibroblastos, que participam no tecido da pele. No entanto, até à data, a relação de sinalização CD109 e STAT3 não foi explorada nestes tipos de células.
CD109 e a sinalização EGFR
receptor do factor de crescimento epidérmico (EGFR) é um membro da família de receptores ErbB. Após a ligação ligante por EGF, EGFR forma dimers, homodímeros ou heterodímeros com outro membro da família ERBB HER2 . Os receptores dimerizados auto-fosforilam-se uns aos outros e depois fosforilam a proteína não-receptora tirosina C-Src cinase, que activa o STAT3 . A ativação do EGFR promove a migração celular, Sobrevivência e proliferação. Em tumores malignos EGFR sobre-expressão está correlacionada com a profundidade da invasão do tumor e ligada ao prognóstico mais pobre . Mutações que levam à sobreexpressão EGFR são detectadas no SCC do pulmão , SCC da cabeça e pescoço e SCC do esófago . O CD109 ancorado com membrana em células SK-MG-1 interage diretamente com o EGFR e reforça a sinalização do FEG, o que aumenta subsequentemente a migração e invasão celular, enquanto o cd109 secretado não tem efeito na sinalização do FEG . EGFR pode mediar os efeitos do CD109 na sinalização do STAT3, o que requer estudos adicionais para elucidar (Fig. 3).
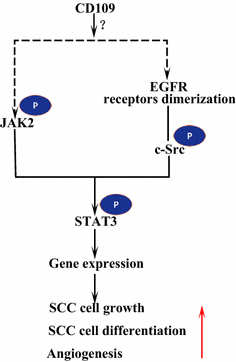
CD109 e sinalização EGFR / STAT3. O CD109 facilita a sinalização do EGFR. Os receptores dimerizados de EGFR fosforilato c-Src, que por sua vez fosforilatos STAT3. CD109 pode também ativa STAT3 através da fosforilação de JAK2, em seguida, promover a SCC crescimento celular, a diferenciação e a angiogênese
A data em que o quadro amplo da tumorigênese e desenvolvimento do SCC permanecem incompletos. Existem conversações cruzadas entre TGF-β e STAT3 ou vias de sinalização EGFR. O CD109 é provavelmente um dos principais efetores da rede de sinalização que regula o SCCs. No entanto, não foram relatadas evidências diretas para delinear os papéis do CD109 na patogênese do CCSC.estudos de amostras de tecidos humanos indicam que o CD109 é altamente expresso em CCSC de múltiplos órgãos, particularmente em células escamosas malignas bem diferenciadas . Através da detecção da expressão CD109 com imunohistoquímica em tecidos humanos, o CD109 pode potencialmente agir como biomarcador para determinar a progressão da CSC. Os estudos actuais sugerem que o CD109 é altamente expresso em CCSC bem diferenciados e que a sua expressão é mais baixa nos CCSC não diferenciados . No entanto, não é claro se o CD109 está associado com invasão vascular, metástase e prognóstico após a cirurgia. Por conseguinte, são necessários mais estudos para explorar o significado clínicoopatológico do CD109 no SCCs numa amostra maior.
CD109 é uma glicoproteína ligada a GPI ,que permite a sua libertação da membrana. A forma solúvel do CD109 também afeta a ligação do TGF-β aos seus receptores, e subsequentemente modular a progressão do SCC . Litvinov et al. descobriu-se que o CD109 libertado da superfície celular para o meio extracelular, e a forma libertada do CD109 mantém a sua capacidade de induzir vias de sinalização intracelulares. Além disso, Sakakura et al. relatou que o CD109 sérico foi libertado pelo tumor xenografado e aumenta proporcionalmente com o volume do xenograft tumoral. Assim, a detecção do nível sérico de CD109 pode ajudar a monitorizar tumores com sobre-expressão de CD109, incluindo SCCs.os exossomas comunicam a lesão tumoral primária e o seu nicho através da sua embalagem contendo proteínas seleccionadas ou outras moléculas . O isolamento e as análises do exossoma associado ao tumor circulante podem servir como biomarcadores para o diagnóstico de pacientes com câncer. O CD109 exógeno foi identificado como um componente do exossoma secretado a partir de 293 células transfectadas, tornando-o um alvo promissor para o diagnóstico exossoma . Ainda assim, até à data, o CD109 não foi relatado em exossomas derivados de CSC ou outras células tumorais. Os mecanismos subjacentes às embalagens CD109 em exossomas merecem uma investigação mais aprofundada.para além do potencial como biomarcador, o CD109 pode ser um alvo para abordagens terapêuticas. CD109 é uma proteína de membrana, que pode ser direcionada diretamente por anticorpos específicos ou enzimas. O CD109 também pode ser reconhecido por veículos-alvo para a entrega de drogas. No entanto, os papéis detalhados do CD109 na patogênese do CCSC ainda não são claros. Por exemplo, é necessário definir a relação entre a expressão mais baixa do CD109 e os CCSC indiferenciados. A intervenção CD109 só pode ser considerada na prática clínica após uma avaliação cuidadosa dos riscos e benefícios.