
pladecellecarcinom (SCC), også kendt som prickle cellecarcinom, er en af de førende årsager til kræftrelateret død i hele verden. SCC er en malign epithelial tumor, der opstår i væv, der giver en barriere mellem en organisme og miljøet, såsom hud, mund, hulrum, spiserør og lunge . På et tidligere tidspunkt ledsages SCC af epidermal keratinisering og sårdannelse i slimhindeoverfladen, og de dybere væv invaderes af SCC ‘ er på et senere tidspunkt. Til mikroskopisk udseende er SCC-cellerne arrangeret i reden, som er omgivet af polygonale celler med forskellige cellegrænser og hyperkromatiske kerner. SCC ‘ er har en høj tendens til at metastasere, normalt gennem regionale lymfeknuder, og kan forårsage systematiske skader på flere organer. Kirurgi, laserterapi og stråling er fortsat den mest anvendte behandling for SCC ‘ er . De unikke cytomorfologiske træk ved SCC-varianter fører til tydelig behandling og resultater .
CELLEOVERFLADEANTIGEN CD109 er et glycosylphosphatidylinositol (GPI)-bundet glycoprotein på cirka 170 kDa og et medlem af familien A2 macroglobulin (a2m) / C3, C4, C5 af thioesterholdige proteiner . Det humane CD109-gen er lokaliseret i kromosom 6K og udgør cirka 3,3% af den samlede CD109-genomiske sekvens med dens 33 eksoner . Den isolerede CD109 cDNA omfatter en 4335 bp åben læseramme, der koder for en 1445 aminosyre (aa) . CD109-proteinet indeholder et 21 aa N-terminal lederpeptid, en formodet agnregion (aa 651-683), et thioesterbindingssted (aa 918-924), en thioesterreaktivitet, der definerer heksapeptid (aa1030–1035) og en C-terminal konsensus GPI-ankersignalsekvens med spaltningen forudsagt at forekomme efter aminosyre 1420 (Fig. 1) . Det N-terminale lederpeptid forankrer proteinet til den indre membran inden i periplasmen og det N-terminale fragment af CD109 udskilt fra celler efter spaltning af furinproteasen . Protease-spaltningen af en stort set uordnet agnregion aktiverer CD109, hvilket resulterer i en konformationsændring, der fælder protease i en burlignende struktur og udsætter den stærkt reaktive thioesterbinding. Thioesterbindingsstedet, som ved aktivering kovalent kan forbinde CD109 med lysinrester på overfladen af den angribende protease. Desuden er den C-terminale konsensus GPI-ankersignalsekvens også kendt som det receptorbindende domæne .

The structure of CD109 protein. Den fulde længde af CD109-protein omfatter 1445 aminosyre (aa), inklusive et 21 aa N-terminalt signalpeptid, et formodet agnområde (aa 651-683), et thioester–bindingssted (aa 918-924), en thioesterreaktivitet, der definerer heksapeptid (aa1030-1035) og en C-terminal konsensus GPI-ankersignalsekvens (aa 1420-1445) med spaltningen forudsagt at forekomme efter aminosyre 1420
cd109 blev først identificeret som et celleoverfladeantigen af et monoklonalt antistof hævet mod den primitive lymfoide/myeloide cellelinie kg1a . Tidligere undersøgelser rapporterede, at CD109 udtrykkes på en undergruppe af føtal og voksen CD34+ positive knoglemarvsmononukleære celler, aktiverede T-lymfoblaster, aktiverede blodplader, endotelceller, mesenkymale stamcelleundersæt og flere humane tumorcellelinjer , men ikke udtrykt i hvilende humane T-celler, blodplader eller perifere blodleukocytter . Især er CD109 blevet påvist i SCC ‘ er i flere organer, herunder lunge , spiserør , livmoderhals , hud , penis , mundhule og galdeblære . CD109 er en komponent i det transformerende vækstfaktor-karr1 (TGF-karr1) receptorsystem og regulerer negativt TGF-karr1 signalering . Tab og gevinst ved funktionsstudier antydede, at CD109 kan være et molekylært mål for udvikling af nye terapeutiske midler til SCC ‘ er af forskellig vævsoprindelse . I denne gennemgang vil vi præsentere en oversigt over den aktuelle viden om CD109 og dens forhold til SCC ‘ er.
CD109-ekspression i SCC ‘er
i tumorvæv blev CD109 immunhistokemisk påvist i SCC’ er såvel som urotelcarcinomer, maligne melanomer , basallignende brystcarcinomer , myksofibrosarcoma , epitel-sarkomer og gliom . Især Shiraki et al. rapporterede CD109-positive perivaskulære tumorceller i humane gliomvæv af lavere kvalitet og i en musemodel rekapituleret humant gliom, hvilket antyder en nøglerolle for CD109 for denne sygdom. Tidligere undersøgelser har vist, at den høje ekspression af CD109 i SCC ‘ er og den begrænsede ekspression i normale pladeceller (tabel 1) . Desuden er CD109 stærkt udtrykt i veldifferentierede SCC ‘er snarere end i moderat eller dårligt differentierede SCC’ er, således er ekspressionsniveauet for CD109 omvendt korreleret med tumorkvalitet .
CD109 og lung SCC
Lung pladecellecarcinom (LSCC) er en af de vigtigste undertyper af ikke-små karcinomer, der er-cellelungekræft (NSCLC) og tegner sig for cirka 20-30% af tilfældene med NSCLC ‘ er . CD109-ekspression blev undersøgt i humane lungecellecarcinomer ved kvantitativ RT-PCR, som viste en signifikant højere ekspression af CD109 i pladecellecarcinomer, men ikke i adenocarcinomer, storcellecarcinomer eller småcellecarcinomer . Derudover Sato et al. brugte et CD109-antistof til at analysere CD109-ekspression i normale lunge-og lungekarcinomvæv. I det normale lungevæv var cd109-ekspression hovedsageligt begrænset til basale celler i den bronchiale og bronchiolære epitel . I lungekarcinomvæv blev der fortrinsvis påvist stærk immunreaktivitet i Lscc ‘ er, men ikke i andre typer lungekarcinomer . Disse resultater tyder på, at CD109 er specifikt opreguleret i Lscc ‘ er.
CD109 og spiserør SCC
spiserør pladecellecarcinom (ESCC) tegner sig for en sjettedel af al kræftrelateret dødelighed, der forekommer ved en højere forekomst i asiatiske lande . Prognosen for ESCC forbliver dårlig, og den 5-årige overlevelsesrate for ESCC er omkring 14-22% . Nylige undersøgelser tyder på, at ændret ekspression af TGF-kursreceptorer bidrager til ESCC-progression, og forhøjet ekspression af hæmmende proteiner af TGF-kurs-signalering korrelerer med dårlig prognose for ESCC . Hashimoto et al. undersøgt CD109-ekspression i ESCC-væv ved kvantitativ RT-PCR, og CD109 mRNA-transkription blev opreguleret i omkring 50% af ESCC ‘ erne. Senere blev proteinniveauet af CD109 i ESCC undersøgt ved immunhistokemi på vævsmikroarrays (TMA) . I det ikke-syge esophageal væv blev CD109-ekspression begrænset i cytosol af de stratificerede epitelceller på et svagt niveau. I alle ESCC-prøver fra TMA ‘ erne viste tråde af ondartede pladepitelceller stærk CD109-farvning i cytosolen. Ingen CD109 positiv farvning blev observeret i andre celletyper. Desuden var CD109 – udtryk højere i godt og moderat differentierede ESCC ‘ er end de dårligt differentierede .
CD109 og livmoderhalsen SCC
livmoderhalskræft (livmoderhalsen) kræft tegner sig for omkring 12% af kræftrelateret død af kvinder . SCC ‘ er tegner sig for 80-85% af alle livmoderhalskræft. Jang et al. viste, at CD109-ekspression var signifikant højere i cervikale SCC ‘ er end i endometrie adenocarcinomer. De undersøgte også CD109-ekspression i fem humane cervikale carcinomcellelinjer og observerede høje niveauer af CD109-ekspression i to SCC-cellelinjer.
CD109 og kutan SCC
kutan pladecellecarcinom (CSCC) er den næst mest almindelige type ikke-melanom hudkræft (NMSC) med en konstant stigende forekomst . Selvom CSCC har en generelt gunstig prognose, er der stadig cirka 1,5–2% af patienterne dør af denne sygdom . CD109-ekspression i CSCCs er blevet undersøgt ved immunhistokemi på TMA ‘ er . I de normale skind blev CD109 svagt udtrykt i det basale lag af epidermale celler, mens tråde af ondartede pladeagtige epidermale celler viste stærk CD109-farvning. Desuden var CD109-udtryk omvendt korreleret med CSCC-kvaliteter. Ligesom ESCC ‘er var udtrykket af CD109 højere i godt og moderat differentierede CSCC’ er end de dårligt differentierede . Penile pladecellecarcinom (PSCC) er en undertype af CSCC ‘ er og er sjældent blevet undersøgt . Dong et al. påvist, at CD109-protein er stærkt udtrykt i maligne pladeceller af PSCC ‘er sammenlignet med normale penile væv på TMA’ er. Desuden valideres ekspressionsmønsteret på friske kirurgiske PSCC-prøver ved immunofluorescens, krt-PCR og vestlig blotting, hvilket antyder, at CD109 kan være en biomarkør for PSCC .
CD109 og oral cavity SCC
Oral pladecellecarcinom (OSCC) tegner sig for 2-3% af alle kræftformer over hele verden . Tab af funktionsmutation i TGF-karrus type II-receptor er en hyppig begivenhed for mundhulen SCC . CD109-ekspression i normale orale væv og OSCC-væv fra 124 patienter blev undersøgt ved immunhistokemisk farvning. Høje niveauer af CD109-ekspression blev ofte påvist i SCC ‘ er og premaligne læsioner i mundhulen, men ikke i normal pladepitel . Desuden var ekspressionsniveauet for CD109 signifikant højere i veldifferentierede OSCCs end i moderat eller dårligt differentierede OSCCs, hvilket indebærer, at CD109-udtryk er korreleret med Differentieringsstadierne af OSCCs . Derudover udviste OSCC-cellelinjer , der overudtrykte CD109, accelereret cellevækst in vitro, hvilket implicerede, at CD109 involverer i progressionen af OSCCs.
CD109 og galdeblære SCC
galdeblærekræft (GBC) er den mest aggressive af galdekræftene med korteste medianoverlevelse . Galdeblære pladecellecarcinom (GSCC) tegner sig for 3% af den ondartede neoplasma i dette organ . Undertyper af gbcs-væv inklusive adenocarcinom (AC), pladecellecarcinom (SCC) og adenoskavamt carcinom (ASC) blev undersøgt på TMA ‘ er ved immunhistokemisk farvning med et CD109-antistof. CD109-farvning var negativ i alle normale galdeblærevæv og AC-væv. I mellemtiden blev CD109-positive celler fundet i 86,7% af SCC ‘er og 91,7% af ASC’ er. Da CD109 tydeligt udtrykkes i ondartede kvadratiske celler i galdeblæren, kan CD109 være en diagnostisk markør for galdeblære SCC ‘er og ASC’ er .
TGF-Lars signalvej og SCCs
TGF-Lars signalvej er involveret i mange cellulære processer, herunder cellevækst, celledifferentiering apoptose og cellulær homeostase . Familien af TGF-β ligander, TGF-β1, TGF-β2 og TGF-β3, binder sig til specifikke transmembrane type i-og type II-serin/threonin-kinase-receptorer (TGF-ßR1 og TGF-ßR2) , hvilket resulterer i aktivering af TGF-ßR1 kinase aktivitet . Den aktiverede TGF-kurrr1 udbreder derefter signalet ved phosphorylering af dets intracellulære substrater, R-SMADs (Smad2 og Smad3) . Smad2 og Smad3 interagerer med TGF-kurrr1 og SARA (Smad-anker til receptoraktivering), et FYVE-domæneprotein, der interagerer direkte med Smad2 og Smad3 , SARA fungerer til at rekruttere Smad2 til TGF-kurr-receptoren, derefter danner de phosphorylerede R-SMADs heteromere komplekser med Co-SMAD (Smad4) . Efter phosphoryleringen og efterfølgende komplekser med Smad4 frigives disse R-Smads-komplekser fra TGF-larr1 og SARA og translokeres derefter ind i kernen, hvor de interagerer med transkriptionsfaktorer , der rekrutterer dem til specifikke promotorelementer af målgener .
Receptorendocytose er en pivotal reguleringsmekanisme i signaltransduktion. TGF-receptorer internaliseres via både clathrin-og caveolae-afhængige veje. Internalisering af de TGF-Kris-receptorer via de clathrin-coatede gruber er blevet forbundet med signalering via Smad2/3 og receptorgenvinding. I modsætning hertil er TGF-release receptor lokalisering i caveolae forbundet med nedregulering af Smad2/3 signalering og receptornedbrydning efter allestedsnærværende af E3-allestedsnærværende ligase Smurf2 . Imidlertid danner hæmmende smad ‘er (Smad6 og Smad7) en særskilt underklasse af SMAD’ er, der virker på en modsat måde til R-smad ‘ er og modvirker signalering . De kan konkurrere med R-smad ‘er om binding til Aktiveret TGF-kurr1 og således hæmme phosphoryleringen af R-smad’ er . Ud over, de rekrutterer E3-allestedsnærværende ligaser til den aktiverede TGF-larr1, hvilket resulterer i receptor allestedsnærværende og nedbrydning, og afslutning af signalering .
dysregulering af TGF-kursvejen er blevet impliceret i flere typer kræft . Undersøgelser har vist, at TGF-Krish-signalering fremkalder en forebyggende virkning i de tidligere stadier af tumorigenese, men en undertrykkende effekt under den senere tumorudvikling . Mutationer i TGF-kurrr1-genet er også fundet i SCC ‘er i huden, hvilket antyder, at inaktivering af TGF-kursist fører til initiering af SCC’ er .
CD109-og TGF-kurssignalvej
CD109 er en TGF-kurssignalreceptor og modulerer TGF-kurssignalreceptoraktivitet på en cellespecifik måde . På celleoverfladen regulerer CD109 negativt TGF-larr1-signalvejen via dannelse af et receptorkompleks med TGF-larr1 og TGF-larr2 i humane keratinocytter . TGF-LARP receptorer nedbrydes efter allestedsnærværende ved E3-allestedsnærværende ligase Smurf2, og internaliseres via både clathrin-afhængige og caveolae-afhængige veje . Biset et al. påvist, at CD109 associeres med caveolin-1 og fremmer TGF-Relas-receptorendocytose. Derudover fremmer CD109 lokalisering af TGF-relo-receptorer i det caveolære rum i nærvær af ligand og letter TGF-relo-receptornedbrydning. CD109 regulerer også lokaliseringen og associeringen af Smad7/Smurf2 med TGF-kurr1. De hæmmende virkninger af CD109 kræver Smad7-ekspression og Smurf2 allestedsnærværende ligaseaktivitet . Endvidere kan CD109 frigives fra celleoverfladen ved hjælp af cellulære lipaser, såsom phosphatidylinositol-specifik phospholipase C (PI-PLC). Den opløselige form af CD109 bevarer sin evne til at binde TGF-krus1 og konfiskere den væk fra TGF-krusreceptorerne .
Vorstenbosch et al. rapporterede, at CD109 differentielt regulerede TGF-kur-inducerede ALK1-Smad1 / 5 versus ALK5-Smad2/3 veje (ALK1 og ALK5 er alle TGF-kurr type i receptorer). De fandt ud af, at TGF-Krust signalering hæmmer endotelcelleproliferation og migration, mens TGF-Krust signalering også inducerer disse processer via ALK1-Smad1/5 . De viser, at ALK1 udtrykkes og lokaliseres sammen med CD109 i musekeratinocytter, og at mus, der overudtrykker CD109 i epidermis, viser forbedret ALK1-Smad1/5 signalering, men nedsat ALK5-Smad2/3 signalering .
desuden er TGF-kurt1 en potent hæmmer af vækst i de fleste epitelceller . Hagivara et al. viste, at orale SCC-cellelinjer overekspression CD109 accelererede celleproliferation og forringede den antiproliferative virkning medieret af TGF-kurt1. I modsætning hertil udviste SCC-celler med CD109-nedslag langsommere cellevækst . Et højt niveau af CD109-ekspression hæmmede Smad2-phosphorylering, således svækket TGF-lar1/Smad2-signalering og forringer TGF-lar1-medieret undertrykkelse af cellevækst, CD109-nedslagning øgede Smad2-phosphorylering ved TGF-larr1-stimulering . Selvom CD109 også regulerer Smad1 / 5 signalering , har den ikke været forbundet med udviklingen af SCC. Tilsammen Letter CD109 udviklingen af SCC ‘ er via hæmning af TGF-kurp-Smad2/3-vejen (Fig. 2).

CD109 og TGF-lys signalering. TGF-Karr binder TGF-karrr2, som rekrutterer og phosphorylerer TGF-karrr1. Aktiveret TGF-kurtr1 udbreder signalet nedstrøms ved direkte phosphorylering af Smad2 og Smad3. Phosphorylerede R-smad ‘ er danner derefter heteromere komplekser med Smad4 kombineret med transkriptionsfaktorer regulerer gentranskription og cellefunktion. TGF-kar-receptorer internaliseres via de clathrin-belagte gruber eller den caveolære vej. CD109 øger TGF-reluris-bindingen til TGF-reluris-receptorer og fremmer TGF-reluris-receptorlokalisering til caveolae, øger i sidste ende TGF-reluris-receptorendocytose og letter TGF-reluris-receptornedbrydning. De inhiberende virkninger af CD109 kræver Smad7-ekspression og Smurf2 allestedsnærværende ligaseaktivitet
CD109 og STAT3-signalering
Signaltransducer og aktivator af transkriptionsfaktor 3 (STAT3) er kritisk for celleproliferation, differentiering, migration, overlevelse og onkogenese . Litvinov et al. rapporteret, at ekspressionen af CD109-protein var markant nedsat i psoriatisk epidermis sammenlignet med tilstødende ikke-involveret hud. CD109 mRNA-ekspression er imidlertid uændret i psoriasisplakker sammenlignet med normal hud, hvilket antyder en mulighed for, at CD109-proteinfrigivelse forbedres i psoriasisk keratinocytter . De foreslog, at frigivet / opløselig CD109 er i stand til at inducere molekylære ændringer, der vides at forekomme i psoriasis . In vitro fandt de, at transfektion af CD109 siRNA nedregulerer STAT3, frigivelse af CD109 fra celleoverfladen af dyrkede humane keratinocytter. Derudover inducerer eksogen/rekombinant CD109 STAT3-signalering i humane keratinocytter . Udover, Chuang et al. rapporterede, at CD109-ekspression var dramatisk opreguleret i metastatiske lungeadenocarcinomceller, og celler, der udtrykte en CD109 shRNA (shCD109), viste en dramatisk reduktion i STAT3-phosphorylering. STAT3-nedslag reducerede kraftigt metastaser, og gendannelse af STAT3-aktivitet øgede shcd109-ekspressive cellers evne til at metastasere . Efter aktivering phosphoryleres STAT3 af ikke-receptor protein tyrosinkinaser janus kinase 2 (JAK2), hvilket fører til dannelsen af STAT3 dimer og translokation i kernen . Inhibering af JAK-kinaseaktivitet i fibroblaster, der overudtrykker CD109, reducerede imidlertid phosphoryleret STAT3 til et niveau svarende til det i forældrecellerne, der udtrykker lave niveauer af CD109, hvilket antyder, at CD109-induceret STAT3-phosphorylering kræver JAK-kinaseaktivitet. Således kan jak / STAT3-signalering formidle virkningerne af CD109 i tumorvækst og metastase .
selvom nedbrydning af CD109 i humane keratinocytter og lungeadenocarcinomceller nedregulerer STAT3-signalering in vitro , viste de CD109-mangelfulde mus modsatte resultater. Mii et al. genererede CD109-mangelfulde mus, som udviste hudabnormiteter inklusive epitelhyperplasi og inflammatorisk celleinfiltration. De rapporterede, at STAT3-phosphorylering i CD109-mangelfulde mus var signifikant højere sammenlignet med vildtypemusene. Derudover er opregulering af STAT3-signalering forbundet med øget proliferation og nedsat differentiering af keratinocytter .
uoverensstemmelsen mellem resultaterne fra in vitro-og in vivo-undersøgelser kan være forårsaget af de systematiske ændringer af mikromiljø i væv fra CD109-mangelfulde mus. Tab af CD109 i alle celler i mus kan ændre det subkutane mikromiljø, som aktiverer STAT3-signalering i keratinocytter. Derudover kan CD109 udøve forskellige regulatoriske virkninger i forskellige celletyper, hvilket fører til celletypespecifik modifikation i STAT3-signalering. Foruden keratinocytter udtrykkes CD109 i endotelceller, epitelceller og fibroblaster, som deltager i udgør hudvævet. Til dato er forholdet mellem CD109 og STAT3-signalering imidlertid ikke blevet undersøgt i disse celletyper.
CD109 og EGFR signalering
Epidermal vækstfaktorreceptor (EGFR) er medlem af erbb-familien af receptorer. Efter LIGANDBINDING fra EGF danner EGFR dimerer, enten homodimerer eller heterodimerer med et andet medlem af erbb-familien HER2 . De dimeriserede receptorer Auto-phosphorylerer hinanden og phosphorylerer derefter ikke-receptorproteintyrosin c-Src-kinase, som aktiverer STAT3 . Aktivering af EGFR fremmer cellemigration, overlevelse og proliferation. I ondartede tumorer er EGFR overekspression korreleret med dybden af invasion af tumoren og knyttet til dårligere prognose . Mutationer , der fører til EGFR-overekspression , opdages i lunge SCC, hoved og hals SCC og spiserør SCC . Den membranforankrede CD109 i SK-MG-1-celler interagerer direkte med EGFR og forbedrer EGF-signalering, hvilket efterfølgende øger cellemigration og invasion, mens den udskilte CD109 ikke har nogen effekt på EGF-signalering . EGFR kan formidle virkningerne af CD109 på STAT3-signalering, hvilket kræver yderligere undersøgelser for at belyse (Fig. 3).
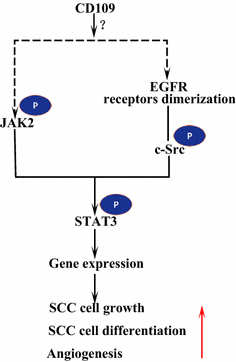
CD109 og EGFR / STAT3 signalering. CD109 Letter EGFR signalering. De dimeriserede EGFR-receptorer phosphorylerer c-Src, som igen phosphorylerer STAT3. CD109 kan også aktivere STAT3 gennem phosphorylering af JAK2 og derefter fremme SCC-cellevækst, differentiering og angiogenese
til dato forbliver det brede billede af tumorigenese og udvikling af SCC ufuldstændig. Der er krydsforhandlinger mellem TGF-Krist og STAT3 eller EGFR signalveje. CD109 er sandsynligvis en af de vigtigste effektorer i signalnetværket, der regulerer SCC ‘ er. Der er dog ikke rapporteret om direkte beviser for at afgrænse cd109 ‘s roller i patogenesen af SCC’ er.
perspektiver
undersøgelser fra humane vævsprøver indikerer, at CD109 er stærkt udtrykt i SCC ‘ er af flere organer, især i veldifferentierede maligne pladeceller . Ved påvisning af CD109-ekspression med immunhistokemi i humane væv kan CD109 potentielt virke som en biomarkør for at bestemme progressionen af SCC. Aktuelle undersøgelser antyder, at CD109 udtrykkes stærkt i veldifferentierede SCC ‘er, og dets udtryk er lavere i ikke-differentierede SCC’ er . Det er imidlertid ikke klart, om CD109 er forbundet med vaskulær invasion, metastase og prognose efter operationen. Derfor er der behov for yderligere undersøgelser for at undersøge den klinikopatologiske Betydning af CD109 i SCC ‘ er i en større prøvestørrelse.
CD109 er et GPI-bundet glycoprotein, som gør det muligt at frigøre det fra membranen . Den opløselige form af CD109 påvirker også bindingen af TGF-Larsen til dets receptorer og modulerer derefter SCC-progression . Litvinov et al. fandt, at CD109 frigivet fra celleoverfladen i det ekstracellulære miljø, og den frigivne form af CD109 bevarer sin evne til at inducere intracellulære signalveje. Udover, Sakakura et al. rapporteret, at serum CD109 blev frigivet af fremmedgjort tumor, og det øges proportionalt med volumenet af tumorgenograft. Derfor kan påvisning af serum CD109-niveau hjælpe med at overvåge tumorer, der overudtrykker CD109 inklusive SCC ‘ er.
eksosomer kommunikerer primær tumorlæsion og dens niche via dens pakke indeholdende udvalgte proteiner eller andre molekyler . Isolering og analyser af cirkulerende tumorassocieret eksosom kan tjene som biomarkører til diagnose af kræftpatienter. Eksogen CD109 er blevet identificeret som en komponent af eksosom udskilt fra transficerede 293 celler, hvilket gør det til et lovende mål for eksosomdiagnose . Indtil videre er CD109 ikke rapporteret i eksosomer afledt af SCC eller andre tumorceller. De underliggende mekanismer for CD109-emballering i eksosomer fortjener mere detaljeret undersøgelse.
ud over potentialet som biomarkør kan CD109 være et mål for terapeutiske tilgange. CD109 er et membranprotein, som kan målrettes direkte af specifikke antistoffer. CD109 kan også genkendes af målrettede køretøjer til levering af narkotika. De detaljerede roller af CD109 i patogenesen af SCC ‘ er er dog stadig uklare. For eksempel skal forholdet mellem lavere udtryk for CD109 og udifferentierede SCC ‘ er defineres. CD109-intervention kan kun overvejes i klinisk praksis, efter at risici og fordele er evalueret omhyggeligt.