
Introduction
nykyinen demografinen muutos länsimaisissa yhteiskunnissa, johon liittyy sekä suhteellinen että absoluuttinen ikäihmisten määrän kasvu, on herättänyt kasvavaa tieteellistä kiinnostusta geriatrisiin kysymyksiin. Tässä yhteydessä onnistuneen vanhenemisen käsitteet ovat yhä tärkeämpiä. Useat tutkimukset ovat osoittaneet, että liikunnalla voi olla keskeinen rooli terveessä ikääntymisessä sekä kognitiivisen heikkenemisen ja hermoston rappeutumissairauksien ehkäisyssä (Colcombe et al., 2006; Erickson et al., 2011). Viimeaikaiset katsaukset, joissa on yhteenveto epidemiologisista, poikkileikkauksista ja interventiaalisista tutkimuksista, tukevat fyysistä aktiivisuutta suotuisana menetelmänä neuroplastisuuden indusoimiseksi myöhäisellä aikuisiällä (Gregory et al., 2012; Voelcker-Rehage and Niemann, 2013; Bamadis et al., 2014). Erickson ym. (2012) ovat todenneet, että korkeampi kardiorespiratorinen kunto ja aerobinen fyysinen aktiivisuus liittyvät suurempiin harmaan aineen tilavuuksiin etuotsalohkoissa ja hippokampuksessa. Kuitenkin paitsi sydän-ja verisuonitauteihin myös koordinoivaan liikuntaan (Niemann et al., 2014)ja kognitiivinen koulutus (Bamadis et al., 2014) on osoitettu indusoivan harmaan aineen plastisuutta ja parantavan kognitiivisia toimintoja vanhemmilla aikuisilla.
eläinkokeissa on esitetty, että liikunnan ja aistinrikastuksen yhdistelmä vaikuttaa voimakkaimmin uusien neuronien syntyyn—pääasiassa hippokampuksessa—ja että vain tämä yhdistelmä takaa vastasyntyneiden solujen kestävän eloonjäämisen (Kempermann ym., 1997; van Praag et al., 2005). Kattenstroth ym. (2013) ovat ehdottaneet, että ”tanssitoimintaa tulisi pitää ihmisen rikastuneiden ympäristöolosuhteiden vastineena, koska ne tarjoavat yksilölle lisääntyneitä aistien, motoristen ja kognitiivisten vaatimusten täyttymyksiä.”Tästä rohkaisevasta lausunnosta huolimatta tutkimuksia tanssiharjoittelun vaikutuksista aivojen rakenteeseen tai toimintaan on vähän. Jälkeen 6 kuukauden tanssi interventio, Kattenstroth et al. (2010) ovat raportoineet merkittäviä parannuksia kognitiivisessa, tunto-ja motorisessa suorituskyvyssä osallistuvilla senioreilla. Prospektiivisen tutkimuksen tulokset, joihin osallistui 469 yli 75-vuotiasta koehenkilöä keskimäärin 5, 1 vuoden seurantajakson aikana, ovat osoittaneet, että tanssimiseen liittyy selvästi pienentynyt dementiariski (Verghese ym., 2003). Kuitenkin, Hüfner et al. (2011) ovat raportoineet vähentyneitä määriä useilla aivoalueilla, mukaan lukien anteriorinen hippokampaalimuodostus ja parieto-Saarelaisen vestibulaarisen aivokuoren osat ammattitanssijoilla ja slacklinereilla verrattuna ei-ammattilaisiin. Tämä havainto voi viitata siihen, että samojen motoristen taitojen intensiivinen ja toistuva harjoittelu johtaa erikoistumisen yhteydessä joidenkin aivoalueiden pienempiin määriin. Tässä tutkimuksessa suunnittelemmekin erityisen tanssiharjoitteluohjelman, joka edellytti osallistujilta jatkuvasti uusien liikemallien opettelua (Müller et al., 2016). Arvioidaksemme tämän toimenpiteen erityishyötyjä vertasimme juuri suunniteltua tanssiohjelmaamme aktiiviseen eikä passiiviseen verrokkiryhmään, joka osallistui perinteiseen kuntoliikuntaohjelmaan, jossa osallistujat tyypillisesti suorittivat toistuvia fyysisiä harjoituksia, kuten pyöräilyä ergometrillä. Lisäksi, koska olimme kiinnostuneita interventioiden ajallisesta dynamiikasta, arvioimme vaikutuksia aivojen rakenteeseen ja toimintaan 6 ja 18 kuukauden harjoittelun jälkeen. Näin pyrimme arvioimaan, onko interventioiden laajentaminen hyödyllistä, koska jotkut aivoalueet saattavat vaatia enemmän harjoittelua kuin toiset, vai onko olemassa raja, jonka jälkeen treenaamisesta tulee haitallista. Lopuksi, etsiessämme mahdollista neuroplastisuuden taustalla olevaa mekanismia, mittasimme ääreisveren BDNF-tasot. Useat tutkimukset ovat osoittaneet, että BDNF edistää uusien neuronien ja synapsien erilaistumista (Huang and Reichardt, 2001; Lessmann and Brigadski, 2009; Park and Poo, 2013; Edelmann et al., 2014). BDNF: ää onkin ehdotettu aikuisen neuroplastisuuden välittäjäksi (Flöel ym., 2010).
materiaalit ja menetelmät
osallistujat ja kokeellinen suunnittelu
tutkimus suunniteltiin 18 kuukauden kontrolloiduksi interventioksi. Magdeburgin Otto-von-Guericken yliopiston eettinen komitea hyväksyi tutkimuksen, ja kaikki tutkittavat allekirjoittivat kirjallisen ilmoituksen, eivätkä he saaneet osallistumisestaan maksua. Tutkimusta varten seulottiin 62 tervettä vanhusta (63-80-vuotiaita), jotka oli rekrytoitu paikallislehtien ilmoitusten kautta. Poissulkukriteereinä olivat klaustrofobia, tinnitus, metalli-implantit, tatuoinnit, diabetes mellitus, masennus (Beck-Depressions Inventory, BDI-II > 13), kognitiiviset vajeet (Mini-Mental State Examination, MMSE < 27), neurologiset sairaudet ja säännöllinen liikunta (≥1 h/viikko). Näiden kriteerien perusteella 10 koehenkilöä jätettiin tutkimuksen ulkopuolelle. Loput 52 osallistujaa satunnaistettiin sitten joko tanssi – tai urheiluryhmään verkkosivujen kautta www.randomization.com ja kontrolloida ikää, MMSE-tilaa ja fyysistä kuntoa. Arvioinnit tehtiin lähtötilanteessa, 6 kuukauden ja 18 kuukauden koulutuksen jälkeen (kuva 1). Kaksikymmentäkaksi osallistujaa suoritti koko intervention ja kaikki mittaukset. Taulukossa 1 on yksityiskohtaiset demografiset tiedot näistä osallistujista. Ryhmäeroja väestötiedoissa ei havaittu.
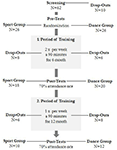
kuva 1. Osallistujien rekrytoinnin vuokaavio.
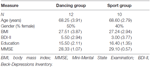
Taulukko 1. Demografiset tiedot osallistujista lähtötilanteessa.
interventiot
interventiot jaettiin kahteen jaksoon. Ensimmäisellä jaksolla koehenkilöt harjoittelivat kahdesti viikossa 90 minuutin sessioissa 6 kuukauden ajan. Käytännön syistä (osallistujien ja kouluttajien saatavuus) toinen 12 kuukauden interventiojakso sisälsi harvennetun kerran viikossa tapahtuvan koulutuksen. Molemmat interventio-ohjelmat suoritettiin ryhmäyhteydessä musiikin kanssa psykososiaalisten vuorovaikutusten hallitsemiseksi. Ehdollista kuormitusta tutkittiin Karvosen ym.mukaan kirjaamalla pulssiarvot harjoituksissa ja laskemalla yksilöllinen treenisyke. (1957) kertoimella 0,6 mittavaan kestävyysharjoitteluun. Näin ollen nämä kaksi harjoitusohjelmaa olivat intensiteetiltään, kestoltaan ja toistotiheydeltään vertailukelpoisia. Molempia valvoivat kokeneet ohjaajat.
tanssiryhmä
tanssiryhmän osallistujat osallistuivat juuri suunniteltuun harjoitusohjelmaan, jossa heitä pyydettiin jatkuvasti opettelemaan uusia liikesarjoja. Nämä koreografiat edellyttivät eri ruumiinosien (eli jalkojen, käsivarsien, rungon) koordinointia avaruudessa erilaisissa rasitusolosuhteissa (fyysinen rasitus, tarkkuus, tilanne ja aikapaine). Koehenkilöiden oli opittava koreografiat ulkoa, mikä asetti suuria vaatimuksia myös muistille. Ohjelmassa oli viisi eri genreä (rivitanssi, jazztanssi, rock ”n” roll ja neliötanssi), jotka vaihtuivat joka neljännen session jälkeen. Intervention aikana koordinaatiovaatimuksia ja aikapainetta lisättiin ottamalla käyttöön monimutkaisempia tanssiliikkeitä ja koreografioita sekä lisäämällä musiikin lyöntejä minuutissa.
Urheiluryhmä
urheiluryhmään osallistuneet suorittivat tavanomaisen kestävyysharjoitteluohjelman, jossa harjoiteltiin lähinnä toistuvia harjoituksia ja jossa koko kehon koordinaatio-ja muistivaatimukset olivat alhaiset. Jokainen istunto koostui 20 minuutin yksiköistä kestävyys -, voima-kestävyys-ja joustoharjoittelua. Kestävyysharjoittelu suoritettiin cycle ergometereillä. Voima-kestävyysyksikössä suoritettiin vaihtelevia liikkeitä (esimerkiksi hauiskiharat, kyykky, istumaannousu), mutta monimutkaisia kokovartaloliikkeitä vältettiin koordinaatiovaatimusten pitämiseksi alhaisina. Notkeusyksikkö koostui pääasiassa venyttelyharjoituksista.
Tulosmittarit
kardiovaskulaarinen kunto
kardiovaskulaarinen kunto arvioitiin fyysisellä työkyky 130-testillä (PwC 130). PWC 130 on teho (watteina mitattuna), jonka koehenkilö pystyy saavuttamaan syketaajuudella 130 bpm. Laskelmissa käytettiin leposykettä ja suhteellista fyysistä kapasiteettia .
neuropsykologinen testaus
tutkittaville tehtiin laaja joukko neuropsykologisia testejä. Tässä tutkimuksessa raportoidaan vain verbaalisen lyhyen ja pitkän aikavälin muistitestin tulokset, ”VLMT” (mukautettu saksankielinen versio ”Rey Auditory Verbal Learning Test”; Helmstaedter and Durwen, 1990) ja tarkkaavaisuustestin akku (test of Attentional Performance (TAP); Zimmermann and FIMM, 2002).
BDNF
Paastoverinäytteet otettiin neuropsykologisten arviointien aamuisin. Verinäytteistä BDNF: n plasmapitoisuudet määritettiin sandwich ELISA-menetelmällä (BDNF DuoSets; r&D Systems, Wiesbaden, Saksa) edellä kuvatulla tavalla (Schega et al., 2016).
MK
MR-kuvat hankittiin 3 Teslan Siemens MAGNETOM Verioon (Syngo MR B17) käyttäen 32-kanavaista pääkäämiä. Korkean resoluution T1-painotetut MPRAGE-sekvenssit hankittiin käyttämällä 3D-magnetoinnissa valmistettua nopeaa gradienttikaikukuvausprotokollaa (224 sagittaaliviipaletta, voxelin koko: 0,8 × 0,8 × 0,8 mm3, TR: 2500 ms, TE: 3,47 ms, TI: 1100 ms, kääntökulma: 7°). MR-kuvat analysoitiin voxel – pohjaisella morfometrialla (VBM), joka toteutettiin SPM 12: ssa (welcome Department of Cognitive Neurology, Lontoo, UK). VBM on koko aivojen puolueeton tekniikka alueellisen harmaan aineen tilavuuden ja kudosten muutosten analysointiin (Ashburner and Friston, 2000).
esikäsittely sisälsi harmaan aineen segmentoinnin, templaatin luomisen Dartelin kautta, spatiaalisen normalisoinnin standardoituun Montrealin neurologiseen instituuttiin (MNI) ja silottamisen Gaussin ytimellä, joka oli 8 mm täysi leveys maksimissa (FWHM).
ryhmien välisten harmaan aineen tilavuusmuutosten erojen analysoimiseksi käytettiin kokonaistekijäryhmää (Tanssi, Urheilu) ja aikaa (0, 6 ja 18 kuukautta). Jos kyseessä on merkittävä ryhmä × aika-vuorovaikutus, peräkkäisten aikaparien väliset post hoc-t-testit (0 vs. 6; 0 vs. 18; 6 vs. 18 kk) laskettiin erikseen kullekin ryhmälle. Kaikkiin analyyseihin sovellettiin raja-arvoa p < 0, 001 (korjaamaton).
tulokset
tulokset esitetään seuraavasti. Etsimme ensin yleistä interventiovaikutusta (tekijäaika), sitten kahden Intervention differentiaalivaikutuksia (vuorovaikutusryhmä × aika); lopuksi tarkempi analyysi ajallisesta dynamiikasta tehtiin post hoc-parivertailuilla.
kardiovaskulaarinen kunto
kardiovaskulaarinen kunto PwC 130-menetelmällä mitattuna (Taulukko 2, suhteellinen fyysinen kapasiteetti) ei eronnut lähtötilanteessa ryhmien välillä. PWC 130-pisteet eivät myöskään nousseet merkittävästi kummankaan toimenpiteen aikana.

Taulukko 2. Välineet ja (SD) kunto, kognitiivinen toiminta, BDNF plasmapitoisuudet ja harmaan aineen kokonaismäärä harjoitusryhmissä yli intervention.
neuropsykologiset testit
merkittävä ajan päävaikutus havaittiin verbaalisessa lyhytkestoisessa muistissa (VLMT early recall; F(2,19) = 6, 438, p = 0, 004, η2 = 0, 253), verbaalisessa pitkäaikaisessa vapaassa muistissa (VLMT late recall; F(2, 19) = 3, 387, P = 0, 049, η2 = 0, 244), verbaalisessa pitkäaikaisessa muistissa (vlmt late recall; F(2, 19) = 3, 387, P = 0, 049, η2 = 0, 244) term recognition (vlmt recognition; F(2,19) = 5, 352, P=0, 009, η2 = 0, 220) ja attention reaction time (Subtest notification; F (2,19) = 19, 156, p < 0, 001, η2 = 0, 489). Merkittäviä aika × ryhmän yhteisvaikutuksia ei ilmennyt.
Post hoc-parivertailuissa havaittiin merkittävää paranemista lähtötilanteesta 18 kuukauteen ja 6 kuukaudesta 18 kuukauteen kaikissa kolmessa VLMT-alaluokassa molemmissa ryhmissä. TAP-reaktioaikojen osalta havaittiin merkittävää paranemista vertailtaessa lähtötilannetta 18 kuukauden tietoihin molemmissa ryhmissä.
BDNF
BDNF: n plasmapitoisuudet analysoitiin verinäytteistä ennen koulutuksen aloittamista sekä 6 kuukauden ja 18 kuukauden kuluttua koulutuksen aloittamisesta. Absoluuttiset BDNF-pitoisuudet plasmassa on esitetty taulukossa 2. BDNF-tason sisäiset muutokset paljastivat tanssiryhmässä merkittävän nousun, kun taas BDNF-taso pysyi tasaisena 6 kuukauden harjoittelun jälkeen lajiryhmässä. BDNF: ssä ei tapahtunut muita muutoksia 18 kuukauden jälkeen missään ryhmässä (kuva 2).
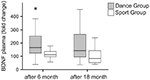
kuva 2. BDNF: n plasmapitoisuuksien yksilökohtaiset muutokset intervention jälkeen. BDNF: n plasmapitoisuudet analysoitiin sellaisten osallistujien verinäytteistä, jotka suorittivat tanssiharjoitusohjelmaa tai urheiluharjoitusohjelmaa ennen harjoittelun aloittamista, 6 kuukauden harjoittelun jälkeen ja 18 kuukauden harjoittelun jälkeen. BDNF-tasojen suhteellinen nousu määritettiin. BDNF-arvot nousivat merkittävästi tanssiryhmässä 6 kuukauden harjoittelun jälkeen (Mann-Whitney U-test, p < 0, 004) ja laskivat lähes hoitoa edeltävän arvon tasolle 18 kuukauden tanssiharjoittelun jälkeen. Urheiluryhmässä (Friedmanin testi, p = 0,319) ei tapahtunut muutosta koko ajan, kun taas tanssiryhmässä (Friedmanin testi, p = 0,028) havaittiin merkittävä muutos koko ajan. Ruutukaavat: minimi, 25. prosenttipiste, mediaani, 75. prosenttipiste. * P ≤ 0, 05.
MK
merkitsevä ryhmä × aika-interaktio havaittiin vasemmassa prekentraalisessa gyruksessa ja oikeassa parahippokampuksessa. Post hoc-t-testit lähtötason ja 6 kuukauden välillä osoittivat harmaan aineen määrän lisääntyneen merkittävästi vain tanssijoiden vasemmassa precentral gyruksessa. Lähtötilanteen ja 18 kuukauden aineiston vertailussa precentral gyruksen lisäksi tanssijoilla näkyi merkittävä harmaan aineen tilavuuden kasvu oikeassa parahippokampuksessa, joka oli myös ainoa merkittävä muutos välissä 6-18 kuukautta. Näin ollen prekentraalisen gyruksen volyymin kasvu ilmeni 6 kuukauden kuluttua ja pysyi vakaana jäljellä olevan tanssiharjoitusvälin ajan, kun taas parahippokampuksen gyruksen muutos tapahtui vasta myöhemmän harjoitusvälin aikana (kuva 3).
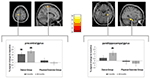
kuva 3. Aika ryhmävuoroanalyysillä, testaamalla suurempia volyymimuutoksia tanssissa verrattuna urheiluryhmään. Merkittävä harmaan aineen lisääntyminen havaittiin precentral gyrus (Montrealin neurologinen instituutti (MNI)-koordinaatit: x = -16; y = -18; z = 77) ja parahippokampuksen gyrus gyrus (MNI-koordinaatit: x = 34; y = -26; z = -20). Laatikkokaaviot osoittavat vokselin huipun suhteellisen harmaan aineen muutokset. * P ≤ 0, 05.
Keskustelu
tässä tutkimuksessa vertailimme joko tanssiohjelmaan tai tavanomaiseen kuntoliikuntaohjelmaan osallistumisen vaikutuksia aivojen toimintaan ja volyymiin terveillä senioreilla. Tanssiohjelma oli vastikään suunniteltu interventio, joka vaati jatkuvasti uusien tanssikoreografioiden opettelua. Perinteinen urheiluohjelma keskittyi lähinnä toistuviin motorisiin harjoituksiin. Huomattavimpana havaintona havaitsimme, että 6 kuukauden harjoittelun jälkeen tanssijoiden vasemman precentral gyruksen volyymit olivat kasvaneet enemmän kuin urheiluryhmässä. Toisen 12 kuukauden harjoittelun jälkeen tanssijoiden oikeassa parahippokampuksessa havaittiin volyymin lisäystä. BDNF-arvot nousivat ensimmäisen 6 kuukauden tanssiharjoittelun aikana ja palautuivat hoitoa edeltäneisiin arvoihin 18 kuukauden jälkeen. Tavanomaisten urheilulajien ryhmässä vastaavaa BDNF: n kasvua ei ollut havaittavissa. Koska sydän-ja verisuonitautien kunto pysyi molemmissa ryhmissä tasaisena toimenpiteiden aikana, havaitut vaikutukset eivät johtuneet fyysisen kunnon parantumisesta vaan näyttivät liittyvän tanssiohjelman erityispiirteisiin. Näitä ominaisuuksia olivat vaatimus jatkuvasti oppia uusia koreografioita (eli muisti), integroida moniaistista tietoa, koordinoida koko kehoa ja navigoida avaruudessa.
aivojen muutokset
precentral gyrus on välttämätön vapaaehtoisten motoristen toimintojen säätelylle. Harmaan aineen tilavuuden kasvu esisentraaligyruksessa tanssiryhmässä saattoikin perustua monimutkaisiin ja alati muuttuviin liikekuvioihin, joita tanssijat joutuivat suorittamaan. Nämä liikkeet vaativat useiden kehon osien samanaikaista koordinointia eri suuntiin ja mukautumista musiikin vaihteleviin rytmeihin (polysentrinen ja polyrytminen). Pohditaan näitä monimutkaisia koordinointivaatimuksia, Brown et al. (2006) ovat raportoineet tanssin aiheuttamista aktivaatioista putamenissa, primaarimotorisessa aivokuoressa ja täydentävässä motorisessa alueella (SMA), kuten Pet osoittaa. Muut tutkimukset ovat osoittaneet yhteyden koordinointivaatimusten (esim., tasapainotus, jongleeraus) ja neuroplastisuus precentral alueella (Boyke et al., 2008; Taubert ym., 2010). Näin ollen tanssiin liittyvä volyymin kasvu tällä alueella vastasi kirjallisuuteen perustuvia odotuksia.
parahippokampaalinen gyrus on osa limbisen järjestelmän ulkokaarta ja sillä on tärkeä rooli työmuistissa ja episodisessa muistinetsinnässä (Pantel ym., 2003). Bliss and Lomon (1973) mukaan parahippokampuksen gyrus muodostaa rajapinnan muistin ja nykyhetken kokemuksellisen tietoisuuden välillä, koska se on yhteydessä otsalohkon alueisiin, jotka liittyvät työmuistiin, ja hippokampukseen, episodisen muistin koodauksen ja avaruudellisen navigoinnin keskeiseen rakenteeseen. Monet VBM-tutkimukset ovat raportoineet ikään liittyvästä volyymin menetyksestä parahippokampaalisilla alueilla (Tisserand et al., 2002). Lisäksi Echávarri et al. (2011) ovat esittäneet, että parahippokampaalinen atrofia on AD: n varhainen biomarkkeri. Havainto siitä, että tanssiohjelmaan osallistuminen pitemmäksi aikaa voi aiheuttaa neuroplastisia prosesseja tällä tärkeällä muistialueella, joten se on erityisen rohkaiseva ehkäisystrategioiden kehittämisen kannalta.
harmaan aineen aivojen plastisuuden ajallinen dynamiikka
havaittu tilavuus kasvaa kahdella eri aikoina kehittyneellä aivoalueella. Tanssi johti volyymin kasvuun Moottorin alueilla 6 kuukauden kuluttua (Katso myös Rehfeld et al., 2016), joka pysyi vakaana seuraavat 12 kuukautta. Tanssiin liittyvä parahippokampuksen volyymin kasvu ilmeni myöhemmin ja havaittiin vasta 18 kuukauden aineistossa. Kahden aivoalueen evoluution aikadynamiikka voi liittyä taustalla olevien solumekanismien eroihin. Eläintutkimus on ehdottanut angiogeneesiä ja uusien dendriittien muodostumista (Thomas et al., 2012) tapahtuvat nopeasti, kun taas neuropiilin muutokset tapahtuvat paljon hitaammin (Black et al., 1990). Ihmisillä on havaittu harmaan aineen määrän nopeaa kasvua etuotsalohkoissa vain 2 viikon motorisen oppimisen jälkeen nuoremmilla aikuisilla (Taubert et al., 2010). Neuroplastisuuden indusoimiseksi hippokampuksessa on suositeltu pidempiä harjoitusjaksoja (Erickson et al., 2012; Niemann et al., 2014).
mielenkiintoista on, että edellä mainitut volyymin lisäykset tutkimuksestamme ovat ristiriidassa Hänggi et al-tutkimuksen tulosten kanssa. (2010), joka paljasti, että ammattimaiset balettitanssijat ovat vähentäneet harmaan aineen määriä vasemmassa premotorisessa aivokuoressa, SMA: ssa, Putamenissa ja superior frontal gyruksessa, ja tulokset Hüfner et al. (2011), joka raportoi vähentyneitä määriä useilla aivoalueilla, mukaan lukien anteriorinen hippokampaalinen muodostuminen ammattitanssijoilla ja slacklinereilla verrattuna ei-ammattilaisiin. Nämä tutkimukset olivat kuitenkin poikkileikkauksellisia havaintotutkimuksia, joissa verrattiin ammattitanssijoiden aivoja ei-ammattilaisten aivoihin. Lisäksi tanssiryhmämme koostui vanhoista noviiseista. On osoitettu, että uuden taidon oppiminen johtaa ensin ylimääräisten hermoresurssien rekrytointiin. Myöhemmin, kun taito muuttuu automaattisemmaksi, tarvitaan vähemmän hermoresursseja, mikä voi johtaa volyymin laskuun pitkäaikaistyöttömillä. Näiden alkuolosuhteiden (aloittelijat, vanhemmat aikuiset) ja erityisen suunnitellun tanssiharjoitteluohjelmamme perusteella, joka edellytti jatkuvasti uutta oppimista liikemalleista, on mahdollista, että erikoistumisen aiheuttamat volyymivaikutukset vähenevät, kuten Hänggi et al. (2010) ei ole havaittu.
yleisesti ajatellaan, että motorinen harjoittelu aiheuttaa aluksi aivojen tilavuuden kasvua. Pitkittynyt harjoittelu johtaa kuitenkin automaatioon, jolla voi olla päinvastaisia vaikutuksia aivokuoren tilavuuteen, koska aivokuoren hallintaa tarvitaan vähemmän, kun motoriset taidot ovat täysin vakiintuneet (Taube, 2008). Tanssiharjoituksemme oli siksi suunniteltu erityisesti välttämään tällaista automaatiota, mikä saattaa selittää sen, miksi ainakaan 18 kuukauden kuluessa tutkimuksessamme ei havaittu aivokuoren volyymin laskua.
kognitiivisissa toiminnoissa ilmeni myös epälineaarista kehitystä, jolloin verbaalinen muisti lisääntyi vasta toisella harjoitusjaksolla. Huomion suorituskyvyn osalta merkittäviä parannuksia havaittiin vain 6 kuukauden kuluttua molemmissa ryhmissä (Rehfeld et al., 2016). Nämä havainnot tukevat aiempia raportteja fysikaalisten toimenpiteiden hyödyllisistä vaikutuksista neuropsykologisiin testeihin (Bamadis et al., 2014). Kognitiivisten kykyjen tiedoissa ei kuitenkaan ilmennyt ryhmäeroja toisin kuin aivodatassa. Toiset ovat raportoineet yhdistetyn kognitiivisen ja fyysisen koulutuksen parempia vaikutuksia verrattuna yksittäisiin interventioihin (Oswald et al., 2006). Laajennamme interventioitamme edelleen testataksemme, voivatko ryhmien väliset erot kognitiossa tulla esiin vielä myöhempinä ajankohtina.
taustalla olevat harmaan aineen plastisuuden solu-ja molekyylimekanismit
vaikka VBM on kuvantamismodaliteetti, joka paljastaa aivojen tilavuusmuutokset, tämä tekniikka ei mahdollista syy-seurausperusteisia johtopäätöksiä taustalla olevista neurofysiologisista prosesseista. Neurogeneesi, synaptogeneesi ja angiogeneesi ovat vain muutamia niistä mekanismeista, joiden on ehdotettu olevan aivojen tilavuusmuutosten perusta (Zatorre et al., 2012). Sydän-ja verisuonikuntoisuuden vaikutusten välittäjinä aivoissa tutkitaan kasvutekijöitä, kuten BDNF, insuliinin kaltainen kasvutekijä (IGF) ja hermokasvutekijä (NGF) (Kirk-Sanchez and McGough, 2014). Kuitenkin meidän tutkimuksessa, toisin kuin aiemmat (Erickson et al., 2011; Maass et al., 2015), ryhmien välillä ei ollut eroja sydän-ja verisuonitautikuntoisuudessa, eivätkä kuntotasot muuttuneet interventioiden aikana. Jälkimmäinen havainto liittyi todennäköisesti yksittäisen sydämen taajuuden säätelyymme, jota pyrimme ylläpitämään aerobisella vyöhykkeellä. BDNF: n muutokset ovat kuitenkin liittyneet myös liikuntaan, sosiaaliseen vuorovaikutukseen ja positiiviseen stressiin (Mattson, 2008), eivätkä kaikki tutkimukset ole havainneet BDNF: n lisääntymistä sydän-ja verisuonitautiharjoittelun jälkeen (Vital et al., 2014). Lopuksi eläinkokeet ovat ehdottaneet, että koordinaatio, mutta ei kestävyysharjoittelu indusoi synaptogeneesiä ja gliasolujen muutoksia(Black et al., 1990). Yhdessä nimetyt lisätekijät, jotka ajavat BDNF: n eritystä, ovat saattaneet olla ratkaisevampia tanssin aikana kuin kuntoilun aikana, mikä selittää, miksi vain tanssijat osoittivat BDNF: n kasvua ensimmäisen 6 kuukauden aikana. Havainto siitä, että BDNF-tasot palautuivat lähtötasolle seuraavien 12 kuukauden aikana, kun parahippokampuksessa havaittiin samanaikaisesti volyymin kasvua, osoittaa kuitenkin, että aikuisten aivojen plastisuuteen täytyy liittyä muitakin tekijöitä kuin ääreisveren BDNF-pitoisuudet.
harjoitetun plastisuuden neurobiologisista mekanismeista myös aivoreservin käsite (Satz et al., 2011) olisi harkittava. Aivoreservin käsite kuvaa yksilöllisiä eroja lisääntyneessä lähtötilanteen adaptiivisessa neuroplastisuudessa, joka tarjoaa suuremman dynaamisen kapasiteetin cortical Circuitsin muokkaamiseen eri stressitekijöille (Barulli and Stern, 2013; Freret et al., 2015).
Perspectives
tutkimuksemme tulokset viittaavat siihen, että pitkäaikainen tanssiva interventio voisi olla toistuvaa liikuntaa parempi neuroplastisuuden indusoinnissa ikääntyvissä ihmisaivoissa. Oletamme, että tämä etu liittyy tanssin multimodaaliseen luonteeseen, jossa yhdistyvät fyysiset, kognitiiviset ja koordinatiiviset haasteet. Tietääksemme tämä on ensimmäinen Pitkittäinen, satunnaistettu tutkimus, jossa suositellaan tanssiohjelmia harmaan aineen ja kognitiivisen heikkenemisen ehkäisemiseksi vanhuksilla. Lisätutkimuksia tarvitaan selventämään tarkemmin tanssin aiheuttaman neuroplastisuuden ajallista dynamiikkaa ja taustalla olevia neurobiologisia mekanismeja sekä sitä, onko tällä toimenpiteellä todella mahdollista vähentää hermoston rappeutumissairauksien, kuten Alzheimerin, riskiä.
Tekijäosuudet
PM suunnitteli ja suoritti tutkimuksen, analysoi tiedot, kirjoitti artikkelin. KR suunnitteli ja toteutti tutkimuksen. MS analysoi tiedot. Ah ja VL suunnittelivat tutkimuksen. MD: tiedonkeruu, artikkelien tarkistus. TB ja JK suunnittelivat tutkimuksen ja analysoivat tiedot. NGM suunnitteli tutkimuksen ja kirjoitti artikkelin.
Eturistiriitalausunto
kirjoittajat toteavat, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, joita voitaisiin pitää mahdollisena eturistiriitana.
Ashburner, J., ja Friston, K. J. (2000). Voxel – pohjainen morfometria-menetelmät. Neuroimage 11, 805-821. doi: 10.1006 / nimg.2000.0582
PubMed Abstract | CrossRef Full Text/Google Scholar
Bamadis, P. D., Vivas, A. B., Styliadis, C., Frantzidis, M., Klados, M., Schlee, W., et al. (2014). Katsaus fyysisiin ja kognitiivisiin interventioihin ikääntymisessä. Neurotieteilijä. Biobehav. Rev. 44, 206-220. doi: 10.1016 / J.neubiorev.2014.03.019
PubMed Abstract | CrossRef Full Text/Google Scholar
Barulli, D., and Stern, Y. (2013). Tehokkuus, kapasiteetti, kompensointi, huolto, plastisuus: syntymässä käsitteitä cognitice reserve. Trendit Cogn. Sci. 17, 502–509. doi: 10.1016 / j.tics.2013.08.012
PubMed Abstract | CrossRef Full Text/Google Scholar
Black, J. E., Isaacs, K. R., Anderson, B. J., Alcantara, A. A., and Greenough, W. T. (1990). Oppiminen aiheuttaa synaptogeneesiä, kun taas motorinen toiminta aiheuttaa angiogeneesiä aikuisten rottien pikkuaivokuoressa. Proc. Natl. Acad. Sci. U S A 87, 5568-5572. doi: 10.1073 / pnas.87.14.5568
PubMed Abstract | CrossRef Full Text/Google Scholar
Bliss, T. V., and Lomo, T. (1973). Aneastetisoidun kanin hammasproteesin synaptisen transmission pitkäkestoinen voimistuminen sen jälkeen, kun perfororeittiä on stimuloitu. J. Fysioli. 232, 331–356. doi: 10.1113 / jphysioli.1973.sp010273
PubMed Abstract | CrossRef Full Text/Google Scholar
Boyke, J., Driemeyer, J., Gaser, C., Büchel, C., and May, A. (2008). Koulutuksen aiheuttama aivojen rakenne muuttuu vanhuksilla. J. Neurotutkija. 28, 7031–7035. doi: 10.1523 / JNEUROSCI.0742-08.2008
PubMed Abstract | CrossRef Full Text/Google Scholar
Brown, S., Martinez, M., and Parsons, M. (2006). Ihmisen tanssin neuronaalinen perusta. Cereb. Cortex 16, 1157-1167. doi: 10.1093/cercor/bhj057
PubMed Abstract | CrossRef Full Text/Google Scholar
Colcombe, S. J., Erickson, K. I., Scalf, P. E., Kim, J. S., Prakash, R., McAuley, E., et al. (2006). Aerobinen kuntoharjoittelu lisää ikääntyvällä ihmisellä aivojen tilavuutta. J. Gerontol. A. Biol. Sci. Med. Sci. 61, 1166–1170. doi: 10.1093 / gerona / 61.11.1166
PubMed Abstract | CrossRef Full Text | Google Scholar
Echávarri, C., Aalten, P., Uyling, H. B. M., Jacobs, H. I. L., Visser, P. J., Gronenschild, M., et al. (2011). Parahippokampuksen surkastuminen Alzheimerin taudin varhaisena biomarkkerina. Aivorakenne. Funct. 215, 265–271. doi: 10.1007 / s00429-010-0283-8
PubMed Abstract | CrossRef Full Text/Google Scholar
Edelmann, E., Lessmann, V., and Brigadski, T. (2014). Pre-ja postsynaptiset käänteet BDNF: n erityksessä ja toiminta synaptisessa plastisuudessa. Neuropharmacology 76, 610-627. doi: 10.1016 / j.neuropharm.2013.05.043
PubMed Abstract | CrossRef Full Text/Google Scholar
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., et al. (2011). Kuntoharjoittelu kasvattaa hippokampuksen kokoa ja parantaa muistia. Proc. Natl. Acad. Sci. U S A 108, 3017-3022. doi: 10.1073 / pnas.1015950108
PubMed Abstract | CrossRef Full Text/Google Scholar
Erickson, K. I., Weinstein, A. M., and Lopez, O. L. (2012). Liikunta, aivojen plastisuus ja Alzheimerin tauti. Kaari. Med. Res. 43, 615-621. doi: 10.1016 / J.arcmed.2012.09.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Flöel, A., Ruscheweyh, R., Krüger, K., Willemer, C., Winter, B., Völker, K., et al. (2010). Liikunta ja muistitoiminnot: ovatko neurotrofiinit ja aivojen harmaan aineen tilavuus puuttuva lenkki? Neuroimage 49, 2756-2763. doi: 10.1016 / j.neuroimage.2009.10.043
PubMed Abstract | CrossRef Full Text/Google Scholar
Freret, T., Gaudreau, P., Schumann-Bard, P., Billard, J. M., and Popa-Wagner, A. (2015). Aivojen varauksen neuroprotektiivisen vaikutuksen taustalla olevat mekanismit myöhäisen elämän masennusta vastaan. J. Neural. Trans. (Wien) 122, S55-S61. doi: 10.1007 / s00702-013-1154-2
PubMed Abstract | CrossRef Full Text/Google Scholar
Gregory, S. M., Parker, B., and Thompson, P. D. (2012). Liikunta, kognitiiviset toiminnot ja aivoterveys: mikä on liikuntaharjoittelun rooli dementian ehkäisyssä? Brain Sci. 2, 684–708. doi: 10.3390 / brainsci2040684
PubMed Abstract | CrossRef Full Text/Google Scholar
Hänggi, J., Koeneke, S., Bezzola, L., and Jäncke, L. (2010). Rakenteellinen neuroplastisuus ammattimaisten naisbalettitanssijoiden sensorimotorisessa verkostossa. Hum. Aivomapp. 31, 1196–1206. doi: 10.1002 / hbm.20928
PubMed Abstract | CrossRef Full Text/Google Scholar
Helmstaedter, C., and Durwen, H. F. (1990). Verbaalinen oppimis-ja retentiotesti. Hyödyllinen ja eriytetty työkalu verbaalisen muistin suorituskyvyn arvioinnissa. Schweiz. Kaari. Neurol. Psychiatr. (1985) 141, 21–30.
PubMed Abstract/Google Scholar
Huang, E. J., and Reichardt, L. F. (2001). Neurotrophins:roolit neuronaalisen kehityksen ja toiminnan. Annu. Pastori Neurotutkija. 24, 677–736. doi: 10.1146 / annurev.neuro.24.1.677
PubMed Abstract / CrossRef Full Text/Google Scholar
Hüfner, K., Binetti, C., Hamilton, D. A., Stephan, T., Flanagin, V. L., Linn, J., et al. (2011). Hippokampuksen muodostumisen rakenteellinen ja toiminnallinen plastisuus ammattitanssijoilla ja slacklinereilla. Hippokampus 21, 855-865. doi: 10.1002 / hipo.20801
PubMed Abstract / CrossRef Full Text / Google Scholar
Karvonen, M. J., Kentala, E., and Mustala, O. (1957). The effects of training on heart rate: a longitudinal study. Ann. Med. Käyt.viim. Biol. Fenn. 35, 307–315.
PubMed Abstract/Google Scholar
Kattenstroth, J. C., Kalisch, T., Holt, S., Tegethoff, M., and dinse, H. R. (2013). Kuuden kuukauden tanssi-interventio parantaa posturaalista, sensorimotorista ja kognitiivista suorituskykyä vanhuksilla vaikuttamatta sydän-ja hengitystoimintoihin. Edessä. Ikääntyvä Neurotutkija. 5:5. doi: 10.3389/fnagi.2013.00005
PubMed Abstract / CrossRef Full Text / Google Scholar
Kattenstroth, J. C., Kolankowska, I., Kalisch, T., ja Dinse, H. R. (2010). Erinomainen sensorinen, motorinen ja kognitiivinen suorituskyky iäkkäillä yksilöillä, joilla on monivuotinen tanssitoiminta. Edessä. Ikääntyvä Neurotutkija. 2:31. doi: 10.3389/fnagi.2010.00031
PubMed Abstract | CrossRef Full Text/Google Scholar
Kempermann, G., Kuhn, H. G., and Gage, F. H. (1997). Enemmän hippokampaalisia neuroneja aikuisilla hiirillä, jotka elävät rikastetussa ympäristössä. Luonto. 386, 493–495. doi: 10.1038/386493a0
PubMed Abstract | CrossRef Full Text | Google Scholar
Kirk-Sanchez, N. J., and McGough, E. L. (2014). Liikunta ja kognitiivinen suorituskyky vanhuksilla: ajankohtaisia näkökulmia. Clin. Interv. Ikääntyminen 9, 51-62. doi: 10.2147 / CIA.S39506
PubMed Abstract | CrossRef Full Text/Google Scholar
Lessmann, V., and Brigadski, T. (2009). Synaptisen BDNF: n erityksen mekanismit, sijainnit ja kinetiikka: päivitys. Neurotieteilijä. 65, 11-22. doi: 10.1016 / J.neures.2009.06.004
PubMed Abstract | CrossRef Full Text/Google Scholar
Maass, A., Düzel, S., Goerke, M., Becke, A., Sobieray, U., Neumann, K., et al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M., Hökelmann, A., Kaufmann, J., et al. (2016). Tanssi-ja terveysliikuntaharjoittelun vaikutukset terveiden senioreiden harmaaseen aivoaineeseen. Sportwiss. 46, 213-222. doi: 10.1007 / s12662-016-0411-6
CrossRef Full Text/Google Scholar
Niemann, C., Godde, B., and Voelcker-Rehage, C. (2014). Sydän-ja verisuonitautien lisäksi myös koordinoitu liikunta lisää hippokampuksen määrää vanhemmilla aikuisilla. Edessä. Ikääntyvä Neurotutkija. 6:170. doi: 10.3389/fnagi.2014.00170
PubMed Abstract / CrossRef Full Text / Google Scholar
Oswald, W. D., Gunzelmann, T., Rupprecht, R., and Hagen, B. (2006). Differential effects of single vs. combined cognitive and physical training with older adults: the SimA study in a 5-year perspective. Euro. J. Ageing 3, 179-192. doi: 10.1007 / s10433-006-0035-z
CrossRef Full Text/Google Scholar
Pantel, J., Kratz, B., Essig, M., and Schröde, J. (2003). Parahippokampuksen volyymivajeet ikääntymiseen liittyvästä kognitiivisesta heikkenemisestä kärsivillä. On. J. Psychiatry 160, 379-382. doi: 10.1176 / appi.ajp.160.2.379
PubMed Abstract / CrossRef Full Text / Google Scholar
Park, H., Ja Kakka, M. M. (2013). Neurotrofiini säätely neuropiirin kehitystä ja toimintaa. Nat. Pastori Neurotutkija. 14, 7–23. doi: 10.1038/nrn3379
PubMed Abstract | CrossRef Full Text | Google Scholar
Rehfeld, K., Hökelmann, A., Kaufmann, J., and Müller, N. G. (2016). ”Effects of a dance vs. a fitness training on brain plasticity, balance performance and attention in healthy seniores: a new approach with SPM 12 for pairwise longitudinal group comparison,” in Proceeding-International Conference of Sports and Neuroscience (Magdeburg), 127-138.
Satz, P., Cole, M. A., Hardy, D. J., and Rassovsky, Y. (2011). Brain and cognitive reserve: mediator (s) and construct validity, a critique. J. Clin. Käyt.viim. Neuropsykolia. 33, 121–130. doi: 10.1080 / 13803395.2010.493151
PubMed Abstract | CrossRef Full Text | Google Scholar
Schega, L., Peter, B., Brigadski, T., Leßmann, V., Isermann, B., Hamacher, D., et al. (2016). Jaksottaisen normobaarisen hypoksian vaikutus aerobiseen kapasiteettiin ja kognitiivisiin toimintoihin iäkkäillä ihmisillä. J. Sci. Med. Sport 19, 941-945. doi: 10.1016 / j.jsams.2016.02.012
PubMed Abstract | CrossRef Full Text/Google Scholar
Taube, W. (2008). Neurofysiologiset mukautukset vastauksena tasapainoharjoitteluun. Dtsch. Z. Sportmed. 63, 273–277. doi: 10.5960 / dzsm.2012.030
CrossRef Full Text / Google Scholar
Taubert, M., Draganski, B., Anwander, A., Müller, K., Horstmann, A., Villringer, A., et al. (2010). Ihmisen aivojen rakenteen dynaamiset ominaisuudet: oppimiseen liittyvät muutokset aivokuoren alueilla ja niihin liittyvät kuituyhteydet. J. Neurotutkija. 30, 11670–11677. doi: 10.1523 / JNEUROSCI.2567-10.2010
PubMed Abstract | CrossRef Full Text/Google Scholar
Thomas, A. G., Dennis, A., Bandettini, P. A., and Johansen-Berg, H. (2012). Aerobisen aktiivisuuden vaikutukset aivojen rakenteeseen. Edessä. Psychol. 3:86. doi: 10.3389 / fpsyg.2012.00086
PubMed Abstract / CrossRef Full Text/Google Scholar
Tisserand, D. J., Pruessner, J. C., Sanz Arigita, E. J., van Boxtel, M. P., Evans, A. C., Jolles, J., et al. (2002). Alueelliset frontaalikuoren tilavuudet pienenevät eri tavoin vanhenemisessa: MRI-tutkimus, jossa verrataan volumetrisiä lähestymistapoja ja vokselipohjaista morfometriaa. Neuroimage 17, 657-669. doi: 10.1016/s1053-8119(02)91173-0
PubMed Abstract | CrossRef Full Text | Google Scholar
van Praag, H., Shubert, T., Zhao, C., and Gage, F. H. (2005). Liikunta tehostaa ikääntyneiden hiirten oppimista ja hippokampaalista neurogeneesiä. J. Neurotutkija. 25, 8680–8685. doi: 10.1523 / JNEUROSCI.1731-05. 2005
PubMed Abstract / CrossRef Full Text/Google Scholar
Verghese, J., Lipton, R. B., Katz, M. J., Hall, C. B., Dervy, C. A., Kuslansky, G., et al. (2003). Vapaa-ajan toiminta ja iäkkäiden dementiariski. N. Engl. J. Med. 348, 2508–2516. doi: 10.1056/NEJMoa022252
PubMed Abstract | CrossRef Full Text/Google Scholar
Vital, T. M., Stein, A. M., de Melo Coelho, F. G., Arantes, F. J., Teodorov, E., Santos-Galduróz, R. F., et al. (2014). Liikunta ja verisuonten endoteelikasvutekijä (VEGF) vanhuksilla: systemaattinen katsaus. Kaari. Gerontolia. Geriatr. 59, 234–239. doi: 10.1016 / J.archger.2014.04.011
PubMed Abstract / CrossRef Full Text / Google Scholar
Voelcker-Rehage, C., ja Niemann, C. (2013). Eri liikuntatyyppeihin liittyvät rakenteelliset ja toiminnalliset aivomuutokset koko eliniän ajan. Neurotieteilijä. Biobehav. Rev. 37, 2268-2295. doi: 10.1016 / J.neubiorev.2013.01.028
PubMed Abstract | CrossRef Full Text/Google Scholar
Zatorre, R. J., Fields, R. D., and Johansen-Berg, H. (2012). Plastisuus harmaa ja valkoinen: neuroimaging muutoksia aivojen rakenteessa oppimisen aikana. Nat. Neurotieteilijä. 15, 528–536. doi: 10.1038 / nn.3045
PubMed Abstract / CrossRef Full Text/Google Scholar
Zimmermann, P., and FIMM, B. (2002). Testi akku huomiota testaus (TAP), versio 1.7, manuaalinen—Part1. Würselen: Psytest.