
Das Plattenepithelkarzinom (SCC), auch bekannt als Stachelzellkarzinom, ist eine der Hauptursachen für krebsbedingte Todesfälle in weltweit. SCC ist ein bösartiger epithelialer Tumor , der in Geweben entsteht, die eine Barriere zwischen einem Organismus und der Umwelt bilden, wie Haut, Mund, Höhle, Speiseröhre und Lunge . In einem früheren Stadium geht SCC mit einer epidermalen Keratinisierung und Ulkusbildung in der Schleimhautoberfläche einher, und in die tieferen Gewebe werden zu einem späteren Zeitpunkt SCCs eingedrungen. Für das mikroskopische Erscheinungsbild sind die SCC-Zellen in Nestern angeordnet, die von polygonalen Zellen mit deutlichen Zellrändern und hyperchromatischen Kernen umgeben sind. SCCs haben eine hohe Tendenz zur Metastasierung, in der Regel durch regionale Lymphknoten, und können systematische Schäden an mehreren Organen verursachen. Chirurgie, Lasertherapie und Bestrahlung sind nach wie vor die am weitesten verbreitete Behandlung für SCCs . Die einzigartigen zytomorphologischen Merkmale von SCC-Varianten führen zu unterschiedlichen Behandlungen und Ergebnissen .Das Zelloberflächenantigen CD109 ist ein Glykosylphosphatidylinositol (GPI) -gebundenes Glykoprotein von etwa 170 kDa und ein Mitglied der a2-Makroglobulin (a2M) / C3, C4, C5-Familie thioesterhaltiger Proteine . Das humane CD109-Gen befindet sich im Chromosom 6q und macht mit seinen 33 Exons etwa 3,3% der gesamten CD109-Genomsequenz aus . Die isolierte CD109 cDNA umfasst einen 4335 bp offenen Leserahmen, der für eine 1445 Aminosäure (aa) kodiert. Das CD109-Protein enthält ein 21 aa N-terminales Leader-Peptid, eine mutmaßliche Bait–Region (aa 651-683), eine Thioester-Bindungsstelle (aa 918-924), ein Thioester-Reaktivitäts-definierendes Hexapeptid (aa1030-1035) und eine C-terminale Konsensus-GPI-Anker-Signalsequenz mit der Spaltung, die nach Aminosäure 1420 vorhergesagt wird (Fig. 1) . Das N-terminale Leader-Peptid verankert das Protein an der inneren Membran innerhalb des Periplasmas und das N-terminale Fragment von CD109, das nach Spaltung durch die Furin-Protease aus Zellen ausgeschieden wird. Die Proteasespaltung einer weitgehend ungeordneten Köderregion aktiviert CD109, was zu einer Konformationsänderung führt, die die Protease in einer käfigartigen Struktur einfängt und die hochreaktive Thioesterbindung freilegt. Die Thioester-Bindungsstelle, die bei Aktivierung CD109 kovalent mit Lysinresten auf der Oberfläche der angreifenden Protease verknüpfen kann. Außerdem ist die C-terminale Konsensus-GPI-Anker-Signalsequenz auch als rezeptor-bindende Domäne bekannt.

The structure of CD109 protein. Die vollständige Länge des CD109-Proteins umfasst 1445 Aminosäure (aa), einschließlich eines 21 aa N-terminalen Signalpeptids, einer mutmaßlichen Köderregion (aa 651-683), einer Thioester–Bindungsstelle (aa 918-924), einer Thioester-reaktivitätsdefinierenden Hexapeptid (aa1030-1035) und einer C-terminalen Konsensus-GPI-Anker-Signalsequenz (aa 1420-1445) mit der Spaltung, die vorhergesagt wird, dass sie nach Aminosäuresequenz auftritt 1420
CD109 wurde zuerst als Zelloberflächenantigen durch einen monoklonalen Antikörper identifiziert, der gegen die primitive lymphoide / myeloische Zelllinie KG1a angehoben wurde . Frühere Studien berichteten, dass CD109 auf einer Untergruppe von fetalen und adulten CD34 + -positiven mononukleären Knochenmarkszellen, aktivierten T-Lymphoblasten, aktivierten Blutplättchen, Endothelzellen, mesenchymalen Stammzelluntergruppen und mehreren menschlichen Tumorzelllinien exprimiert wird , jedoch nicht in ruhenden menschlichen T-Zellen, Blutplättchen oder peripheren Blutleukozyten exprimiert wird . Insbesondere wurde CD109 in SCCs mehrerer Organe nachgewiesen, darunter Lunge, Speiseröhre, Gebärmutterhals , Haut , Penis, Mundhöhle und Gallenblase . CD109 ist eine Komponente des transformierenden Wachstumsfaktor-β1 (TGF-β1) -Rezeptorsystems und reguliert die TGF-β1-Signalisierung negativ . Verlust und Funktionsgewinn Studien deuteten darauf hin, dass CD109 ein molekulares Ziel für die Entwicklung neuer Therapeutika für SCCs verschiedener Gewebeherkunft sein könnte . In dieser Übersicht werden wir eine Zusammenfassung des aktuellen Wissens über CD109 und seine Beziehung zu SCCs präsentieren.
CD109-Expression in SCCs
In Tumorgeweben wurde CD109 immunhistochemisch sowohl in SCCs als auch in Urothelkarzinomen, malignen Melanomen, basalähnlichen Mammakarzinomen, Myxofibrosarkomen, Epithelsarkomen und Gliomen nachgewiesen . Insbesondere Shiraki et al. berichtete CD109-positive perivaskuläre Tumorzellen in menschlichen niedriggradigen Gliomgeweben und in einem Mausmodell rekapitulierten menschliches Gliom, was auf eine Schlüsselrolle von CD109 für diese Krankheit hindeutet. Frühere Studien haben gezeigt, dass die hohe Expression von CD109 in SCCs und die begrenzte Expression in normalen Plattenepithelzellen (Tabelle 1) . Darüber hinaus wird CD109 eher in gut differenzierten SCCs als in mäßig oder schlecht differenzierten SCCs stark exprimiert, so dass das Expressionsniveau von CD109 umgekehrt mit dem Tumorgrad korreliert .
CD109 und Lungen-SCC
Das Lungen-Plattenepithelkarzinom (LSCC) ist einer der wichtigsten Subtypen von nicht-kleinzelligem Lungenkrebs (NKL NSCLC) und macht etwa 20-30% der Fälle von NSCLC aus . Die CD109-Expression wurde in humanen Lungenzellkarzinomen durch quantitative RT-PCR untersucht, die eine signifikant höhere Expression von CD109 in Plattenepithelkarzinomen zeigte, nicht jedoch in Adenokarzinomen, großzelligen Karzinomen oder kleinzelligen Karzinomen . Darüber hinaus haben Sato et al. verwendete einen CD109-Antikörper zur Analyse der CD109-Expression in normalen Lungen- und Lungenkarzinomgeweben. In den normalen Lungengeweben war die CD109-Expression hauptsächlich auf Basalzellen der Bronchial- und Bronchiolarepithelien beschränkt . In Lungenkarzinomgeweben wurde eine starke Immunreaktivität bevorzugt in LSCCs nachgewiesen, jedoch nicht in anderen Arten von Lungenkarzinomen . Diese Ergebnisse deuten darauf hin, dass CD109 in LSCCs spezifisch hochreguliert ist.
CD109 und Ösophagus-SCC
Das Plattenepithelkarzinom der Speiseröhre (ESCC) macht ein Sechstel aller krebsbedingten Mortalität aus und tritt in asiatischen Ländern häufiger auf . Die Prognose von ESCC bleibt schlecht und die 5-Jahres-Überlebensrate von ESCC beträgt etwa 14-22% . Neuere Studien deuten darauf hin, dass eine veränderte Expression von TGF-β-Rezeptoren zur ESCC-Progression beiträgt, und eine erhöhte Expression von inhibitorischen Proteinen der TGF-β-Signalisierung korreliert mit einer schlechten Prognose von ESCC . Hashimoto et al. untersuchte die CD109-Expression in ESCC-Geweben durch quantitative RT-PCR, und die CD109-mRNA-Transkription wurde in etwa 50% der ESCCs hochreguliert. Später wurde der Proteinspiegel von CD109 in ESCC durch Immunhistochemie an Gewebemikroarrays (TMA) untersucht . Im nicht erkrankten Ösophagusgewebe war die CD109-Expression im Cytosol der geschichteten Epithelzellen auf einem schwachen Niveau eingeschränkt. In allen ESCC-Proben aus den TMAs zeigten Stränge maligner Plattenepithelzellen eine starke CD109-Färbung im Cytosol. Bei anderen Zelltypen wurde keine CD109-positive Färbung beobachtet. Darüber hinaus war die CD109-Expression in gut und mäßig differenzierten ESCCs höher als in schlecht differenzierten .
CD109 und Gebärmutterhalskrebs SCC
Gebärmutterhalskrebs (Gebärmutterhalskrebs) macht etwa 12% des krebsbedingten Todes von Frauen aus . SCCs machen 80-85% aller Gebärmutterhalskrebserkrankungen aus . Zhang et al. zeigte, dass die CD109-Expression in zervikalen SCCs signifikant höher war als in endometrialen Adenokarzinomen. Sie untersuchten auch die CD109-Expression in fünf humanen Zervixkarzinom-Zelllinien und beobachteten hohe CD109-Expression in zwei SCC-Zelllinien.
CD109 und kutanes SCC
Das kutane Plattenepithelkarzinom (CSCC) ist die zweithäufigste Form von Nicht-Melanom-Hautkrebs (NMSC) mit stetig steigender Inzidenz . Obwohl CSCC eine allgemein günstige Prognose haben, gibt es immer noch etwa 1,5–2% der Patienten an dieser Krankheit sterben . Die CD109-Expression in CSCCs wurde durch Immunhistochemie an TMAs untersucht . In der normalen Haut wurde CD109 in der Basalschicht der Epidermiszellen schwach exprimiert, während Stränge maligner plattenepithelialer Epidermiszellen eine starke CD109-Färbung aufwiesen. Außerdem war die CD109-Expression umgekehrt mit den CSCC-Werten korreliert. Wie ESCCs war die Expression von CD109 in gut und mäßig differenzierten CSCCs höher als in schlecht differenzierten . Das Plattenepithelkarzinom des Penis (PSCC) ist ein Subtyp von CSCCs und wurde selten untersucht . Dong et al. gezeigt, dass CD109-Protein in malignen Plattenepithelzellen von PSCCs im Vergleich zu normalen Penisgeweben auf TMAs stark exprimiert wird. Darüber hinaus wird das Expressionsmuster an frischen chirurgischen PSCC-Proben durch Immunfluoreszenz, qRT-PCR und Western-Blotting validiert, was darauf hindeutet, dass CD109 ein Biomarker für PSCC sein kann .
CD109 und Mundhöhle SCC
Orales Plattenepithelkarzinom (OSCC) macht 2-3% aller Krebserkrankungen weltweit aus . Funktionsverlust Mutation im TGF-β-Typ-II-Rezeptor ist ein häufiges Ereignis für Mundhöhle SCC . Die CD109-Expression in normalen oralen Geweben und OSCC-Geweben von 124 Patienten wurde durch immunhistochemische Färbung untersucht. Hohe CD109-Expression wurden häufig in SCCs und prämalignen Läsionen der Mundhöhle nachgewiesen, jedoch nicht in normalen Plattenepithelien . Darüber hinaus war das Expressionsniveau von CD109 in gut differenzierten OSCCs signifikant höher als in mäßig oder schlecht differenzierten OSCCs, was impliziert, dass die CD109-Expression mit den Differenzierungsstadien von OSCCs korreliert . Darüber hinaus zeigten OSCC-Zelllinien, die CD109 überexprimierten, in vitro ein beschleunigtes Zellwachstum , was impliziert, dass CD109 am Fortschreiten von OSCCs beteiligt ist.
CD109 und Gallenblase SCC
Gallenblasenkrebs (GBC) ist der aggressivste Gallenkrebs mit dem kürzesten medianen Überleben . Das Plattenepithelkarzinom der Gallenblase (GSCC) macht 3% des malignen Neoplasmas dieses Organs aus . Subtypen von GBCs-Geweben, einschließlich Adenokarzinom (AC), Plattenepithelkarzinom (SCC) und adenosquamöses Karzinom (ASC), wurden an TMAs durch immunhistochemische Färbung mit einem CD109-Antikörper untersucht. Die CD109-Färbung war in allen normalen Gallenblasengeweben und AC-Geweben negativ. Inzwischen wurden CD109-positive Zellen in 86,7% der SCCs und 91,7% der ASCs gefunden. Da CD109 in malignen Plattenepithelzellen in der Gallenblase deutlich exprimiert wird, kann CD109 ein diagnostischer Marker für SCCs und ASCs der Gallenblase sein .
TGF-β-Signalweg und SCCs
Der TGF-β-Signalweg ist an vielen zellulären Prozessen beteiligt, darunter Zellwachstum, Zelldifferenzierungsapoptose und zelluläre Homöostase . Die Familie der TGF-β-Liganden, TGF-β1, TGF-β2 und TGF-β3, bindet an spezifische Transmembran-Typ-I- und Typ-II-Serin / Threonin-Kinase-Rezeptoren (TGF-ßR1 und TGF-ßR2) , was zur Aktivierung der TGF-ßR1-Kinase-Aktivität führt . Das aktivierte TGF-ßR1 propagiert dann das Signal durch Phosphorylierung seiner intrazellulären Substrate, R-SMADs (Smad2 und Smad3) . Smad2 und Smad3 interagieren mit TGF-ßR1 und SARA (Smad-Anker für Rezeptoraktivierung), einem FYVE-Domänenprotein, das direkt mit Smad2 und Smad3 interagiert, SARA rekrutiert Smad2 für den TGF-β-Rezeptor , dann bilden die phosphorylierten R-SMADs heteromere Komplexe mit Co-SMAD (Smad4) . Nach der Phosphorylierung und anschließenden Komplexen mit Smad4 werden diese R-Smads-Komplexe aus TGF-ßR1 und SARA freigesetzt und translozieren dann in den Zellkern, wo sie mit Transkriptionsfaktoren interagieren, die sie für spezifische Promotorelemente von Zielgenen rekrutieren .
Die Rezeptorendozytose ist ein zentraler Regulationsmechanismus bei der Signaltransduktion. TGF-β-Rezeptoren werden sowohl über Clathrin- als auch über Caveolae-abhängige Wege internalisiert. Die Internalisierung der TGF-β-Rezeptoren über die mit Clathrin beschichteten Pits wurde mit der Signalübertragung über Smad2 / 3 und dem Rezeptorrecycling in Verbindung gebracht. Im Gegensatz dazu ist die Lokalisierung des TGF-β-Rezeptors in Caveolen mit einer Herunterregulation der Smad2 / 3-Signalisierung und einem Rezeptorabbau nach Ubiquitinierung durch die E3-Ubiquitin-Ligase Smurf2 verbunden . Inhibitorische Smads (Smad6 und Smad7) bilden jedoch eine eigene Unterklasse von Smads, die entgegengesetzt zu R-Smads wirken und die Signalgebung antagonisieren . Sie können mit R-Smads um die Bindung an aktiviertes TGF-ßR1 konkurrieren und somit die Phosphorylierung von R-Smads hemmen. Darüber hinaus rekrutieren sie E3-Ubiquitin-Ligasen an das aktivierte TGF-ßR1, was zu Rezeptor-Ubiquitinierung und -abbau sowie Beendigung der Signalübertragung führt .
Die Dysregulation des TGF-β-Signalwegs wurde mit mehreren Krebsarten in Verbindung gebracht . Studien haben gezeigt, dass die TGF-β-Signalisierung während der früheren Stadien der Tumorentstehung eine vorbeugende Wirkung hervorruft, aber eine unterdrückende Wirkung während der späteren Tumorentwicklung . Mutationen im TGF-ßR1-Gen wurden auch in SCCs der Haut gefunden, was darauf hindeutet, dass die Inaktivierung von TGF-β zur Initiierung von SCCs führt .
CD109- und TGF-β-Signalweg
CD109 ist ein TGF-β-Co-Rezeptor und moduliert die TGF-β-Signalrezeptoraktivität zellspezifisch . Auf der Zelloberfläche reguliert CD109 den TGF-β1-Signalweg negativ über die Bildung eines Rezeptorkomplexes mit TGF-ßR1 und TGF-ßR2 in menschlichen Keratinozyten . TGF-β-Rezeptoren werden nach Ubiquitinierung durch die E3-Ubiquitin-Ligase Smurf2 abgebaut und sowohl über clathrinabhängige als auch über Caveolae-abhängige Wege internalisiert . Bizet et al. gezeigt, dass CD109 mit Caveolin-1 assoziiert und fördert TGF-β-Rezeptor-Endozytose. Zusätzlich fördert CD109 Lokalisierung der TGF-β Empfänger in das caveolar Fach in Anwesenheit des Ligands und erleichtert Verminderung des TGF-β Empfängers. CD109 reguliert auch die Lokalisierung und die Assoziation von Smad7/Smurf2 mit TGF-ßR1. Die inhibitorischen Wirkungen von CD109 erfordern Smad7-Expression und Smurf2-Ubiquitin-Ligase-Aktivität . Darüber hinaus kann CD109 von der Zelloberfläche durch zelluläre Lipasen wie Phosphatidylinositol-spezifische Phospholipase C (PI-PLC) freigesetzt werden. Die lösliche Form von CD109 behält seine Fähigkeit, TGF-β1 zu binden und es von den TGF-β-Rezeptoren weg zu konfiszieren .
Vorstenbosch et al. berichtet, dass CD109 differentiell reguliert TGF-β-induzierte ALK1-Smad1 / 5 versus ALK5-Smad2 / 3 Wege (ALK1 und ALK5 sind alle TGF-β Typ I Rezeptoren). Sie fanden heraus, dass die TGF-β-Signalisierung die Proliferation und Migration von Endothelzellen hemmt, während die TGF-β-Signalisierung diese Prozesse auch über ALK1-Smad1 / 5 induziert . Sie zeigen, dass ALK1 exprimiert wird und co-lokalisiert mit CD109 in Maus-Keratinozyten und dass Mäuse überexprimieren CD109 in der Epidermis Anzeige verstärkt ALK1-Smad1/5 Signalisierung, aber verringert ALK5-Smad2/3 Signalisierung .
Außerdem ist TGF-β1 ein potenter Wachstumshemmer in den meisten Epithelzellen . Hagiwara et al. gezeigt, dass orale SCC-Zelllinien Überexpression CD109 beschleunigte die Zellproliferation und beeinträchtigte die anti-proliferative Wirkung vermittelt durch TGF-β1. Im Gegensatz dazu zeigten SCC-Zellen mit CD109-Knockdown ein langsameres Zellwachstum . Ein hohes Maß an CD109-Expression inhibierte die Smad2-Phosphorylierung, dämpfte somit die TGF-β1 / Smad2-Signalisierung und beeinträchtigte die TGF-β1-vermittelte Unterdrückung des Zellwachstums, CD109 Knockdown erhöhte die Smad2-Phosphorylierung durch TGF-β1-Stimulation . Obwohl CD109 auch die Smad1 / 5-Signalisierung reguliert , wurde es nicht mit der Entwicklung von SCC in Verbindung gebracht. Zusammen erleichtert CD109 die Entwicklung von SCCs durch Hemmung des TGF-β-Smad2 / 3-Signalwegs (Abb. 2).

CD109- und TGF-β-Signalisierung. TGF-β bindet das TGF-ßR2, das das TGF-ßR1 rekrutiert und phosphoryliert. Aktiviertes TGF-ßR1 propagiert das Signal stromabwärts, indem es Smad2 und Smad3 direkt phosphoryliert. Phosphorylierte R-SMADs bilden dann heteromere Komplexe mit Smad4, kombiniert mit Transkriptionsfaktoren, regulieren die Gentranskription und Zellfunktion. TGF-β-Rezeptoren internalisieren über die mit Clathrin beschichteten Pits oder den Caveolarweg. CD109 erhöht die TGF-β-Bindung an TGF-β-Rezeptoren und fördert die Lokalisierung des TGF-β-Rezeptors in den Hohlräumen, erhöht letztendlich die TGF-β-Rezeptorendozytose und erleichtert den Abbau des TGF-β-Rezeptors. Die inhibitorischen Wirkungen von CD109 erfordern Smad7-Expression und Smurf2-Ubiquitin-Ligase-Aktivität
CD109- und STAT3-Signalisierung
Der Signalwandler und Aktivator des Transkriptionsfaktors 3 (STAT3) ist entscheidend für die Zellproliferation, Differenzierung, Migration, Überleben und Onkogenese . Litwinow et al. es wurde berichtet, dass die Expression des CD109-Proteins in der psoriatischen Epidermis im Vergleich zu benachbarter unbeteiligter Haut deutlich verringert war. Die CD109-mRNA-Expression ist jedoch in psoriatischen Plaques im Vergleich zu normaler Haut unverändert, was auf die Möglichkeit hindeutet, dass die CD109-Proteinfreisetzung in psoriatischen Keratinozyten verstärkt ist . Sie schlugen vor, dass freigesetztes / lösliches CD109 molekulare Veränderungen induzieren kann, von denen bekannt ist, dass sie bei Psoriasis auftreten . In vitro fanden sie heraus, dass die Transfektion von CD109-siRNA die STAT3-Freisetzung von CD109 von der Zelloberfläche kultivierter menschlicher Keratinozyten herunterreguliert. Darüber hinaus induziert exogenes / rekombinantes CD109 die STAT3-Signalisierung in menschlichen Keratinozyten . Außerdem Chuang et al. berichtet, dass die CD109-Expression in metastasierten Lungenadenokarzinomzellen dramatisch hochreguliert war und Zellen, die eine CD109-shRNA (shCD109) exprimierten, eine dramatische Reduktion der STAT3-Phosphorylierung zeigten. STAT3 Knockdown stark reduziert Metastasen, und die Wiederherstellung der STAT3-Aktivität erhöht die Fähigkeit der shCD109-exprimierenden Zellen zu metastasieren . Bei Aktivierung wird STAT3 durch die Nicht-Rezeptorprotein-Tyrosinkinasen Januskinase 2 (JAK2) phosphoryliert, was zur Bildung von STAT3-Dimer und Translokation in den Zellkern führt . Die Hemmung der JAK-Kinaseaktivität in Fibroblasten, die CD109 überexprimieren, reduzierte jedoch das phosphorylierte STAT3 auf ein Niveau, das dem in den elterlichen Zellen ähnlich ist, die niedrige CD109-Spiegel exprimieren, was darauf hindeutet, dass die CD109-induzierte STAT3-Phosphorylierung eine JAK-Kinaseaktivität erfordert. Somit könnte die JAK / STAT3-Signalisierung die Auswirkungen von CD109 auf das Tumorwachstum und die Metastasierung vermitteln .Obwohl der Knockdown von CD109 in menschlichen Keratinozyten und Lungenadenokarzinomzellen die STAT3-Signalisierung in vitro herunterreguliert , zeigten die CD109-defizienten Mäuse entgegengesetzte Ergebnisse. In: Mii et al. erzeugte CD109-defiziente Mäuse, die Hautanomalien einschließlich epithelialer Hyperplasie und entzündlicher Zellinfiltration aufwiesen. Sie berichteten, dass die STAT3-Phosphorylierung in CD109-defizienten Mäusen im Vergleich zu den Wildtyp-Mäusen signifikant höher war. Darüber hinaus ist die Hochregulation der STAT3-Signalisierung mit einer erhöhten Proliferation und einer gestörten Differenzierung von Keratinozyten verbunden .
Die Diskrepanz zwischen den Ergebnissen von In-vitro- und In-vivo-Studien könnte durch die systematischen Veränderungen der Mikroumgebung in den Geweben von CD109-defizienten Mäusen verursacht werden. Der Verlust von CD109 in allen Zellen von Mäusen könnte die subkutane Mikroumgebung verändern, die die STAT3-Signalisierung in Keratinozyten aktiviert. Darüber hinaus kann CD109 unterschiedliche regulatorische Effekte in verschiedenen Zelltypen ausüben, was zu einer zelltypspezifischen Modifikation der STAT3-Signalisierung führt. Zusätzlich zu Keratinozyten wird CD109 in Endothelzellen, Epithelzellen und Fibroblasten exprimiert , die an der Bildung des Hautgewebes beteiligt sind. Bisher wurde die Beziehung zwischen CD109- und STAT3-Signalisierung in diesen Zelltypen jedoch nicht untersucht.
CD109- und EGFR-Signalisierung
Der epidermale Wachstumsfaktor-Rezeptor (EGFR) gehört zur ErbB-Rezeptorfamilie. Bei Ligandenbindung durch EGF bildet EGFR Dimere, entweder Homodimere oder Heterodimere mit einem anderen Mitglied der ErbB-Familie HER2 . Die dimerisierten Rezeptoren phosphorylieren sich gegenseitig und phosphorylieren dann die Nicht-Rezeptorprotein-Tyrosin-c-Src-Kinase, die STAT3 aktiviert . Die Aktivierung von EGFR fördert die Zellmigration, das Überleben und die Proliferation. Bei malignen Tumoren korreliert die EGFR-Überexpression mit der Eindringtiefe des Tumors und ist mit einer schlechteren Prognose verbunden . Mutationen, die zu einer EGFR-Überexpression führen, werden in Lungen-SCC , Kopf-Hals-SCC und Ösophagus-SCC nachgewiesen . Das membranverankerte CD109 in SK-MG-1-Zellen interagiert direkt mit EGFR und verstärkt die EGF-Signalisierung, was anschließend die Zellmigration und -invasion erhöht, während das sekretierte CD109 keinen Einfluss auf die EGF-Signalisierung hat . EGFR könnte die Auswirkungen von CD109 auf die STAT3-Signalisierung vermitteln, was weitere Studien zur Aufklärung erfordert (Abb. 3).
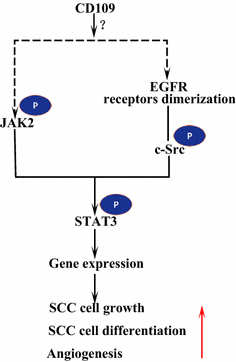
CD109- und EGFR/STAT3-Signalisierung. CD109 erleichtert die EGFR-Signalisierung. Die dimerisierten EGFR-Rezeptoren phosphorylieren c-Src, welches wiederum STAT3 phosphoryliert. CD109 kann auch aktiviert STAT3 durch Phosphorylierung von JAK2, dann fördern SCC Zellwachstum, Differenzierung und Angiogenese
Bis heute das breite Bild der Tumorentstehung und Entwicklung von SCC unvollständig bleiben. Es gibt Kreuzgespräche zwischen TGF-β- und STAT3- oder EGFR-Signalwegen. CD109 ist wahrscheinlich einer der Schlüsseleffektoren des Signalisierungsnetzwerks, das SCCs reguliert. Es wurden jedoch keine direkten Hinweise auf die Rolle von CD109 bei der Pathogenese von SCCs berichtet.
Perspektiven
Studien aus menschlichen Gewebeproben zeigen, dass CD109 in SCCs multipler Organe, insbesondere in gut differenzierten malignen Plattenepithelzellen, stark exprimiert wird . Durch den Nachweis der CD109-Expression mit Immunhistochemie in menschlichen Geweben kann CD109 möglicherweise ein Biomarker sein, um das Fortschreiten von SCC zu bestimmen. Aktuelle Studien legen nahe, dass CD109 in gut differenzierten SCCs stark exprimiert wird und seine Expression in nicht differenzierten SCCs niedriger ist . Es ist jedoch nicht klar, ob CD109 mit Gefäßinvasion, Metastasierung und Prognose nach der Operation assoziiert ist. Daher sind weitere Studien erforderlich, um die klinisch-pathologische Bedeutung von CD109 bei SCCs in einer größeren Stichprobengröße zu untersuchen.
CD109 ist ein GPI-gebundenes Glykoprotein, das es ermöglicht, aus der Membran freigesetzt zu werden . Die lösliche Form von CD109 beeinflusst auch die Bindung von TGF-β an seine Rezeptoren und moduliert anschließend die SCC-Progression . Litwinow et al. gefunden, dass CD109 von der Zelloberfläche in das extrazelluläre Milieu freigesetzt wird, und die freigesetzte Form von CD109 behält seine Fähigkeit, intrazelluläre Signalwege zu induzieren. Außerdem Sakakura et al. berichtet, dass Serum CD109 wurde von xenografted Tumor freigesetzt und es steigt proportional mit dem Volumen des Tumors Xenograft. Daher kann der Nachweis des CD109-Serumspiegels dazu beitragen, Tumoren zu überwachen, die CD109 überexprimieren, einschließlich SCCs.
Exosomen kommunizieren primäre Tumorläsion und ihre Nische über ihr Paket, das ausgewählte Proteine oder andere Moleküle enthält . Die Isolierung und Analyse zirkulierender tumor-assoziierter Exosome kann als Biomarker für die Diagnose von Krebspatienten dienen. Exogenes CD109 wurde als Bestandteil von Exosomen identifiziert, die aus transfizierten 293-Zellen ausgeschieden werden, was es zu einem vielversprechenden Ziel für die Exosomendiagnose macht . Bisher wurde CD109 jedoch nicht in Exosomen berichtet, die von SCC oder anderen Tumorzellen stammen. Die zugrunde liegenden Mechanismen der CD109-Verpackung in Exosomen verdienen eine detailliertere Untersuchung.Neben dem Potenzial als Biomarker könnte CD109 ein Ziel für therapeutische Ansätze sein. CD109 ist ein Membranprotein, das direkt von spezifischen Antikörpern oder Enzymen angegriffen werden kann. CD109 könnte auch von gezielten Vehikeln zur Arzneimittelabgabe erkannt werden. Die detaillierte Rolle von CD109 bei der Pathogenese von SCCs ist jedoch noch unklar. Zum Beispiel muss die Beziehung der niedrigeren Expression von CD109 und undifferenzierten SCCs definiert werden. Eine CD109-Intervention kann in der klinischen Praxis nur in Betracht gezogen werden, nachdem die Risiken und der Nutzen sorgfältig abgewogen wurden.