
introduktion
den nuværende demografiske ændring i vestlige samfund, der involverer både en relativ og en absolut stigning i antallet af ældre, har udløst stigende videnskabelig interesse for geriatriske problemer. I denne sammenhæng bliver begreber om vellykket aldring stadig vigtigere. Flere undersøgelser har vist, at fysisk træning kan spille en nøglerolle i sund aldring og i forebyggelsen af kognitiv tilbagegang og neurodegenerative sygdomme (Colcombe et al., 2006; Erickson et al., 2011). Nylige anmeldelser, der opsummerer epidemiologiske, tværsnits-og interventionsundersøgelser, understøtter fysisk aktivitet som en gunstig metode til at inducere neuroplasticitet i sen voksen alder (Gregory et al., 2012; Voelcker-Rehage og Niemann, 2013; Bamadis et al., 2014). Erickson et al. (2012) har konkluderet, at højere kardiorespiratorisk kondition og aerobe fysiske aktivitetsniveauer er forbundet med større gråstofvolumener i de præfrontale regioner og hippocampus. Men ikke kun kardiovaskulær fitness, men også koordinerende motion (Niemann et al., 2014) og kognitiv træning (Bamadis et al., 2014) har vist sig at inducere grå Stof plasticitet og forbedre kognitive funktioner hos ældre voksne.
dyreforskning har antydet, at en kombination af fysisk træning med sensorisk berigelse har den stærkeste effekt på dannelsen af nye neuroner—overvejende i hippocampus—og at kun denne kombination sikrer den vedvarende overlevelse af de nyfødte celler (Kempermann et al., 1997; van Praag et al., 2005). Kattenstroth et al. (2013) har foreslået, at “danseaktiviteter skal betragtes som et ækvivalent med berigede miljøforhold for mennesker, da de giver et individ øgede sensoriske, motoriske og kognitive krav.”På trods af denne opmuntrende erklæring er undersøgelser, der undersøger effekten af dansetræning på hjernens struktur eller funktion, knappe. Efter en 6-måneders danseintervention, Kattenstroth et al. (2010) har rapporteret betydelige forbedringer i kognitiv, taktil og motorisk ydeevne hos deltagende seniorer. Resultaterne af en prospektiv undersøgelse af 469 forsøgspersoner ældre end 75 år over en median opfølgningsperiode på 5,1 år har indikeret, at dans er forbundet med en markant reduceret risiko for demens (Verghese et al., 2003). Imidlertid, h Kristfner et al. (2011) har rapporteret reducerede mængder i flere hjerneområder, herunder den forreste hippocampale dannelse og dele af parieto-insular vestibulær bark hos professionelle dansere og slackliners sammenlignet med ikke-professionelle. Dette fund kan indikere, at intensiv og gentagen træning af de samme motoriske færdigheder fører i forbindelse med specialisering til reducerede mængder af nogle hjerneområder. I denne undersøgelse designer vi derfor et specielt dansetræningsprogram, der konstant krævede, at deltagerne lærte nye bevægelsesmønstre (m Pristller et al., 2016). At vurdere de specifikke fordele ved denne intervention, vi sammenlignede vores nydesignede danseprogram med en aktiv snarere end en passiv kontrolgruppe, der deltog i et konventionelt sundhedssport-fitnessprogram, hvor deltagerne typisk udførte gentagne fysiske øvelser såsom cykling på et ergometer. Desuden, fordi vi var interesserede i den tidsmæssige dynamik i interventionerne, vurderede vi virkningerne på hjernens struktur og funktion efter 6 og 18 måneders træning. Dermed forsøgte vi at vurdere, om det er fordelagtigt at udvide interventioner, fordi nogle hjerneområder muligvis kræver mere træning end andre, eller om der er en grænse, hvorefter mere træning bliver skadelig. Endelig, på jagt efter en potentiel mekanisme, der ligger til grund for neuroplasticitet, målte vi BDNF-niveauerne i det perifere blod. Flere undersøgelser har antydet, at BDNF fremmer differentieringen af nye neuroner og synapser (Huang og Reichardt, 2001; Lessmann og Brigadski, 2009; Park and Poo, 2013; Edelmann et al., 2014). BDNF er derfor blevet foreslået at være en mediator for voksen neuroplasticitet (FL Kurstel et al., 2010).
materialer og metoder
deltagere og eksperimentelt Design
undersøgelsen blev designet som en 18-måneders kontrolleret intervention. Undersøgelsen blev godkendt af etikudvalget for Otto-von-Guericke University, Magdeburg, og alle emner underskrev en skriftlig informeret, og de modtog ikke betaling for deres deltagelse. Toogtres raske ældre personer (63-80 år) rekrutteret via meddelelser i lokale aviser blev screenet for undersøgelsen. Udelukkelseskriterierne var klaustrofobi, tinnitus, metalimplantater, tatoveringer, diabetes mellitus, depression (Beck-Depressions Inventory, BDI-II > 13), kognitive underskud (Mini-Mental tilstandsundersøgelse, MMSE < 27), neurologiske sygdomme og regelmæssig træning (kg 1 time/uge). På baggrund af disse kriterier blev 10 personer udelukket. De resterende 52 deltagerne blev derefter tilfældigt tildelt enten dansen eller sportsgruppen ved hjælp af hjemmesiden www.randomization.com og kontrol for alder, MMSE status og fysisk kondition. Evalueringer blev udført ved baseline efter 6 og efter 18 måneders træning (Figur 1). Toogtyve deltagere gennemførte hele interventionen og alle målinger. Tabel 1 indeholder detaljerede demografiske data for disse deltagere. Der blev ikke fundet nogen gruppeforskelle med hensyn til de demografiske data.
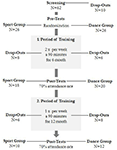
Figur 1. Rutediagram over deltagernes rekruttering.
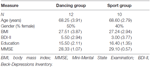
tabel 1. Demografiske oplysninger om deltagerne ved baseline.
interventioner
interventionerne blev opdelt i to perioder. I den første periode trænede fagene to gange om ugen i 90-min sessioner i 6 måneder. Af praktiske grunde (tilgængelighed af deltagere og undervisere) omfattede den anden 12-måneders interventionsperiode en reduceret træningsfrekvens på en gang om ugen. Begge interventionsprogrammer blev udført i en gruppekontekst med musik til kontrol for psykosociale interaktioner. Betinget belastning blev undersøgt ved at registrere pulsværdierne under træningssessionerne og ved at beregne den individuelle træningspuls ifølge Karvonen et al. (1957) med en faktor 0,6 til omfattende udholdenhedstræning. Derfor var de to træningsprogrammer sammenlignelige med hensyn til intensitet, varighed og hyppighed. Begge blev overvåget af erfarne instruktører.
dansegruppe
deltagerne i dansegruppen deltog i et nydesignet træningsprogram, hvor de konstant blev bedt om at lære nye bevægelsessekvenser. Disse koreografier krævede koordinering af forskellige kropsdele (dvs.ben, arme, bagagerum) i rummet under forskellige belastningsforhold (fysisk belastning, præcision, situation og tidspres). Fagene måtte lære koreografierne udenad og stillede således også høje krav til hukommelsen. Programmet bestod af fem forskellige genrer (line dance, rock ” n ” roll og firkantet dans), som blev skiftet efter hver fjerde session. I løbet af interventionen blev koordinerende krav og tidspres øget ved at introducere mere komplekse dansetrin og koreografier og ved at øge beats per minut i musikken.
Sportsgruppe
deltagerne i sportsgruppen gennemførte et konventionelt styrke-udholdenhedstræningsprogram med hovedsageligt gentagne øvelser og lave krav med hensyn til koordinering af hele kroppen og hukommelse. Hver session omfattede 20-min enheder af udholdenhed, styrke-udholdenhed og fleksibilitet træning. Udholdenhedstræningen blev udført på cykel ergometre. I styrke-udholdenhedsenheden blev der udført skiftende bevægelser (f.eks. biceps krøller, knebøjninger, sit-ups), men komplekse hele kropsbevægelser blev undgået for at holde koordinative krav lave. Fleksibilitetsenheden bestod hovedsageligt af strækøvelser.
resultatmål
kardiovaskulær kondition
kardiovaskulær kondition blev vurderet ved den fysiske arbejdskapacitet 130 Test (PVC 130). 130 er den effekt, som et motiv er i stand til at opnå på et cyklusergometer under en puls på 130 bpm. Til beregninger brugte vi hvilepuls og relativ fysisk kapacitet .
neuropsykologisk test
et omfattende batteri af neuropsykologiske tests blev udført på emnerne. Med henblik på den aktuelle undersøgelse rapporteres kun resultaterne af en verbal kort – og langtidshukommelsestest, “VLMT” (en tilpasset tysk version af “Rey Auditory Verbal Learning Test”; Helmstaedter og durven, 1990) og et opmærksomhedstestbatteri (Test of Attentional Performance (TAP); Simmermann og Fimm, 2002).
BDNF
fastende blodprøver blev taget om morgenen af de neuropsykologiske vurderinger. Fra blodprøverne blev plasmakoncentrationerne af BDNF bestemt af smørrebrød Elisas (BDNF DuoSets; R & D Systems, Viesbaden, Tyskland) som tidligere beskrevet (Schega et al., 2016).
MR
MR-billeder blev erhvervet på en 3 Tesla Siemens MAGNETOM Verio (Syngo MR B17) ved hjælp af en 32-kanals hovedspole. T1-vægtede MPRAGESEKVENSER med høj opløsning blev erhvervet ved hjælp af en 3D-magnetiseringspræpareret hurtig gradient-ekko-billeddannelsesprotokol (224 sagittale skiver, vokselstørrelse: 0,8 liter 0,8 liter 0,8 mm3, TR: 2500 ms, TE: 3,47 ms, TI: 1100 ms, vippevinkel: 7 liter). MR-billederne blev analyseret ved hjælp af vokselbaseret morfometri (VBM) implementeret i SPM 12 (velkommen Institut for kognitiv neurologi, London, UK). VBM er en helhjerne upartisk teknik til analyse af regionalt gråstofvolumen og vævsændringer (Ashburner og Friston, 2000).
forbehandling involverede segmentering af grå Stof, oprettelse af skabeloner via DARTEL, rumlig normalisering til standardiseret Montreal Neurological Institute (MNI) rum og udjævning med en Gaussisk kerne på 8 mm fuld bredde ved halv maksimum (FHM).
for at analysere forskellen i gråstofvolumenændringer mellem grupper blev der anvendt et fuldt faktordesign med faktorgruppen (dans, sport) og tid (0, 6 og 18 måneder). I tilfælde af signifikante gruppelys-tidsinteraktioner, post hoc t-test mellem på hinanden følgende par tidspunkter (0 vs. 6; 0 vs. 18; 6 vs. 18 måneder) blev beregnet separat for hver gruppe. En tærskel på p < 0,001 (ukorrigeret) blev anvendt til alle analyser.
resultater
præsentationen af resultaterne er struktureret som følger. Vi kiggede først efter en generel interventionseffekt (faktortid) og kiggede derefter efter differentielle effekter af de to interventioner (interaktionsgruppe kurstid); endelig blev en mere detaljeret analyse af den tidsmæssige dynamik udført via post hoc parvise sammenligninger.
kardiovaskulær kondition
kardiovaskulær kondition målt ved PVC 130 (tabel 2, relativ fysisk kapacitet) var ikke forskellig mellem grupperne ved baseline. Desuden steg PV 130-scoren ikke signifikant i løbet af nogen af interventionerne.

tabel 2. Midler og (SD) til fitness, kognitiv funktion, BDNF plasmaniveauer og total gråstofvolumen inden for træningsgrupper over interventionen.
neuropsykologiske Tests
der blev observeret en signifikant hovedeffekt af tiden for verbal kortvarig hukommelse (VLMT tidlig tilbagekaldelse; F(2,19) = 6,438, p = 0,004, lus2 = 0,253), verbal langvarig fri tilbagekaldelse (vlmt sen tilbagekaldelse; F(2,19) = 3,387, p = 0,049, lus2 = 0,244), verbal langsigtet fri tilbagekaldelse (vlmt sen tilbagekaldelse; F(2,19) = 3,387, p = 0,049, lus2 = 0,244), verbal langvarig tilbagekaldelse genkendelse (vlmt-genkendelse; f(2,19) = 5,352, p=0,009, lus2 = 0,220) og OPMÆRKSOMHEDSREAKTIONSTID (Subtest-fleksibilitet; f (2,19) = 19,156, p < 0,001, lus2 = 0,489). Der opstod ingen signifikant tid, hvor der var interaktion mellem grupper af grupper.
post hoc parvise sammenligninger viste signifikante forbedringer fra baseline til 18 måneder og fra 6 til 18 måneder i alle tre vlmt-underkategorier i begge grupper. Med hensyn til TAP-reaktionstider blev der set en signifikant forbedring i sammenligningen af baseline til 18-måneders data i begge grupper.
BDNF
plasmaniveauer af BDNF blev analyseret i blodprøver inden træningens begyndelse samt 6 måneder og 18 måneder efter træningens begyndelse. Absolutte BDNF-plasmaniveauer er opsummeret i tabel 2. De intraindividuelle ændringer i BDNF-niveau afslørede en markant stigning i dansegruppen, mens det intraindividuelle BDNF-niveau forblev konstant efter 6 måneders træning i sportsgruppen. Ingen yderligere ændringer i BDNF forekom efter 18 måneder i nogen gruppe (figur 2).
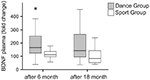
figur 2. Intraindividuelle ændringer i BDNF-plasmaniveauer efter intervention. BDNF plasmaniveauer blev analyseret i blodprøver af deltagere, der udførte et dansetræningsprogram eller et sportstræningsprogram inden træningens begyndelse, efter 6 måneders træning og efter 18 måneders træning. Den relative stigning i BDNF-niveauer blev kvantificeret. BDNF-niveauerne steg markant i dansegruppen efter 6 måneders træning (Mann-Hvidney U-test, p < 0.004) og faldt næsten til baseline for forbehandlingsværdien efter 18 måneders dansetræning. Der var ingen ændring over hele tidsforløbet i sportsgruppen (Friedman test, p = 0.319), mens der blev observeret en signifikant ændring over hele tidsforløbet i dansegruppen (Friedman test, p = 0.028). Boks plots: minimum, 25. percentil, median, 75. percentil. * p 0,05.
MR
der blev observeret en signifikant gruppe-Relas-tidsinteraktion i den venstre precentrale gyrus og den højre parahippocampal. Post hoc t-test mellem baseline-og 6-måneders data viste en signifikant stigning i gråstofvolumen i den venstre precentrale gyrus hos kun danserne. I sammenligningen af baseline og 18-måneders data ud over den precentrale gyrus udviste danserne en betydelig stigning i mængden af grå stof i den rigtige parahippocampale gyrus, hvilket også var den eneste signifikante ændring i intervallet fra 6 til 18 måneder. Således opstod volumenforøgelsen i den precentrale gyrus efter 6 måneder og forblev stabil over det resterende dansetræningsinterval, mens ændringen i parahippocampal gyrus kun forekom under det senere træningsinterval (figur 3).
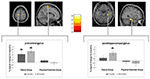
figur 3. Tid efter gruppeinteraktionsanalyse, test for større volumenændringer i dansen sammenlignet med sportsgruppen. En signifikant stigning i grå Stof blev fundet i precentral gyrus (Montreal Neurological Institute (Mni)-koordinater: h = -16; y = -18; h = 77) og i parahippocampal gyrus gyrus (MNI-koordinater: h = 34; y = -26; å = -20). Kasseplottene viser de relative gråstofændringer i topvoksen. * p 0,05.
Diskussion
i denne undersøgelse sammenlignede vi virkningerne af deltagelse i enten et danseprogram eller et konventionelt sportsprogram for fysisk fitness på hjernefunktion og volumen hos raske seniorer. Dansprogrammet var en nydesignet intervention, der krævede konstant at lære nye dansekoreografier. Det konventionelle sportsprogram fokuserede hovedsageligt på gentagne motorøvelser. Som et hovedfund observerede vi, at efter 6 måneders træning var mængderne i dansernes venstre precentrale gyrus steget mere end dem i sportsgruppen. Efter yderligere 12 måneders træning blev der observeret en yderligere volumenforøgelse i dansernes højre parahippocampale gyrus. BDNF-niveauer steg i løbet af de første 6 måneder af dansetræning og vendte tilbage til forbehandlingsværdierne efter 18 måneder. I den konventionelle sportsgruppe var en lignende stigning i BDNF ikke tydelig. Fordi de kardiovaskulære fitnessniveauer i løbet af interventionerne forblev konstante i begge grupper, kunne de observerede effekter ikke tilskrives forbedringer i fysisk kondition, men syntes i stedet at være relateret til de specifikke træk ved dansprogrammet. Disse funktioner omfattede kravet om konstant at lære nye koreografier (dvs.hukommelse), at integrere multisensorisk information, at koordinere hele kroppen og navigere i rummet.
hjerneændringer
den precentrale gyrus er afgørende for styringen af frivillige motorfunktioner. Stigningen i gråstofvolumen i den precentrale gyrus i dansegruppen kan derfor have været baseret på de komplekse og stadigt skiftende bevægelsesmønstre, som danserne måtte udføre. Disse bevægelser krævede samtidig koordinering af flere dele af kroppen i forskellige retninger og tilpasning til musikens forskellige rytmer (polycentrisk og polyrytmisk). Reflektere over disse komplekse koordineringskrav, Brun Et al. (2006) har rapporteret dans-induceret aktiveringer i putamen, den primære motorbark og det supplerende motorområde (SMA), som vist af PET. Andre undersøgelser har indikeret en sammenhæng mellem koordinationskrav (f.eks. balancering, jonglering) og neuroplasticitet i det precentrale område (Boyke et al., 2008; Taubert et al., 2010). Derfor var den danserelaterede volumenforøgelse på dette område i overensstemmelse med forventningerne baseret på litteraturen.
den parahippocampale gyrus er en del af den ydre bue af det limbiske system og spiller en vigtig rolle i arbejdshukommelsen og episodisk hukommelse hentning (Pantel et al., 2003). Ifølge Bliss og Lomo (1973) udgør parahippocampal gyrus grænsefladen mellem hukommelse og nutidens oplevelsesbevidsthed, fordi den er forbundet med perforantkanalen både til regioner i frontallappen, der er forbundet med arbejdshukommelse, og til hippocampus, den centrale struktur i episodisk hukommelseskodning og rumlig navigation. Mange vbm-undersøgelser har rapporteret et aldersrelateret volumentab i parahippocampale regioner (Tisserand et al., 2002). Desuden, Ech Kurvarri et al. (2011) har antydet, at parahippocampal atrofi er en tidlig biomarkør for AD. Observationen om, at deltagelse i et danseprogram i en længere periode kan inducere neuroplastiske processer i denne afgørende hukommelsesregion, derfor især opmuntrende med hensyn til udvikling af forebyggelsesstrategier.
Temporal dynamik af grå Stof hjerne plasticitet
det observerede volumen stiger i de to hjerneområder udviklet på forskellige tidspunkter. Dans førte til en volumenforøgelse i motorområderne efter 6 måneder (Se også Rehfeld et al., 2016), som forblev stabil i de efterfølgende 12 måneder. Den dansassocierede volumenforøgelse i parahippocampal gyrus opstod senere og blev kun observeret i 18-måneders data. De forskellige tidsmæssige dynamikker i udviklingen af de to hjerneområder kan være relateret til forskelle i de underliggende cellulære mekanismer. Dyreforskning har antydet, at angiogenese og dannelsen af nye dendritter (Thomas et al., 2012) forekommer hurtigt, mens ændringer i neuropilen forekommer meget langsommere (Black et al., 1990). Hos mennesker er der observeret hurtige stigninger i gråstofvolumen i de præfrontale regioner efter kun 2 ugers motorisk læring hos yngre voksne (Taubert et al., 2010). For at inducere neuroplasticitet i hippocampus er længere træningsperioder blevet anbefalet (Erickson et al., 2012; Niemann et al., 2014).
interessant nok er de førnævnte volumenforøgelser fra vores undersøgelse i modsætning til resultaterne af H. (2010), som afslørede, at professionelle balletdansere har reduceret mængden af grå stof i venstre premotorbark, SMA, Putamen og overlegen frontal gyrus og resultaterne af H. (2011), som rapporterer reducerede mængder i flere hjerneområder, herunder den forreste hippocampale dannelse hos professionelle dansere og slackliners sammenlignet med ikke-professionelle. Disse undersøgelser var imidlertid tværsnitsobservationsundersøgelser, der sammenlignede professionelle dansers hjerner med ikke-professionelle. Derudover bestod vores dansegruppe af gamle gamle nybegyndere. Det har vist sig, at læring af en ny færdighed først fører til rekruttering af yderligere neurale ressourcer. Senere, når færdigheden bliver mere automatisk, er der behov for mindre neuronale ressourcer, som kan føre til volumenfald hos dem med langvarig erfaring. Baseret på disse indledende betingelser (nybegyndere, ældre voksne) og vores specialdesignede dansetræningsprogram, som krævede konstant ny læring af bevægelsesmønstre, er det muligt, at virkningerne af specialiseringsinduceret volumen falder som rapporteret af H Larsnggi et al. (2010) overholdes ikke.
det menes generelt, at motorisk træning oprindeligt inducerer hjernevolumen øges. Langvarig træning fører imidlertid til automatisering, hvilket kan have de modsatte virkninger på kortikalt volumen, fordi mindre kortikal kontrol er nødvendig, efter at motorens færdigheder er fuldt etableret (Taube, 2008). Vores dansetræningsprogram var derfor specielt designet til at undgå sådan automatisering, hvilket kan forklare, hvorfor der i det mindste inden for 18 måneder ikke blev observeret nogen kortikale volumenfald i vores undersøgelse.
kognitive funktioner viste også ikke-lineær udvikling, hvorved verbal hukommelse kun steg i den anden træningsperiode. Med hensyn til opmærksomhedsydelse var signifikante forbedringer observerbare efter kun 6 måneder i begge grupper (Rehfeld et al., 2016). Disse fund understøtter tidligere rapporter om de gavnlige virkninger af fysiske indgreb på neuropsykologiske tests (Bamadis et al., 2014). I data om kognitiv evne, i modsætning til hjernedataene, opstod der imidlertid ingen gruppeforskelle. Andre har rapporteret overlegne effekter af kombineret kognitiv og fysisk træning i modsætning til enkeltinterventioner., 2006). Vi vil udvide vores interventioner yderligere for at teste, om gruppeforskelle i kognition kan opstå på endnu senere tidspunkter.
underliggende cellulære og molekylære mekanismer for Gråstofplasticitet
selvom VBM er en billeddannelsesmodalitet, der afslører volumenændringer i hjernen, tillader denne teknik ikke kausale konklusioner vedrørende de underliggende neurofysiologiske processer. Neurogenese, synaptogenese og angiogenese er blot nogle få af de mekanismer, der er blevet foreslået at være grundlaget for hjernevolumenændringer., 2012). Som mediatorer af virkningerne af kardiovaskulær fitness på hjernen undersøges vækstfaktorer som BDNF, insulinlignende vækstfaktor (IGF) og nervevækstfaktor (NGF) (Kirk-Sanches og McGough, 2014). Men i vores undersøgelse, i modsætning til tidligere (Erickson et al., 2011; Maass et al., 2015), var der ingen forskelle i kardiovaskulær kondition mellem grupper, og fitnessniveauerne ændrede sig ikke under interventionerne. Sidstnævnte observation var sandsynligvis relateret til vores kontrol af den individuelle hjertefrekvens, som vi havde til formål at opretholde i det aerobe område. Imidlertid har BDNF-ændringer også været forbundet med fysisk aktivitet, social interaktion og positiv stress (Mattson, 2008), og ikke alle undersøgelser har observeret en BDNF-stigning efter kardiovaskulær træning (Vital et al., 2014). Endelig har dyreforskning antydet, at koordinering, men ikke udholdenhedstræning inducerer synaptogenese og glialændringer (Black et al., 1990). Sammen kan de navngivne yderligere faktorer, der driver BDNF-sekretion, have været mere afgørende under Dans end under fitnessaktiviteter, hvilket forklarer, hvorfor kun dansere viste en BDNF-stigning i de første 6 måneder. Observationen af, at BDNF-niveauer vendte tilbage til baseline i de følgende 12 måneder, mens volumenforøgelser samtidig blev observeret i parahippocampal gyrus, indikerer imidlertid, at der skal være andre faktorer involveret i voksen hjerne plasticitet end dem, der er repræsenteret af BDNF-niveauer i det perifere blod.
vedrørende neurobiologiske mekanismer for udøvet induceret plasticitet også begrebet hjernereserve (sat et al., 2011) bør overvejes. Begrebet hjernereserve beskriver individuelle forskelle i en øget baseline adaptiv neuroplasticitet, som giver større dynamisk kapacitet til ombygning af kortikale kredsløb til forskellige stressfaktorer (Barulli og Stern, 2013; Freret et al., 2015).
perspektiver
resultaterne af vores undersøgelse antyder, at en langvarig danseintervention kunne være bedre end gentagen fysisk træning til at inducere neuroplasticitet i den aldrende menneskelige hjerne. Vi antager, at denne fordel er relateret til dansens multimodale karakter, der kombinerer fysiske, kognitive og koordinerende udfordringer. Så vidt vi ved, er dette den første langsgående, randomiserede undersøgelse, der anbefaler danseprogrammer som et middel til at forhindre grå stof og kognitiv tilbagegang hos ældre. Yderligere forskning er nødvendig for at afklare mere detaljeret den tidsmæssige dynamik og de underliggende neurobiologiske mekanismer for dansinduceret neuroplasticitet, og om denne intervention virkelig har potentialet til at reducere risikoen for neurodegenerative sygdomme som f.eks.
Forfatterbidrag
PM designet og udført forskningen, analyserede dataene, skrev artiklen. KR designede og udførte forskningen. MS analyserede dataene. AH og VL designet forskningen. MD: dataindsamling, artikel revision. TB og JK designede forskningen, analyserede dataene. NGM designet forskningen, skrev artiklen.
interessekonflikt Erklæring
forfatterne erklærer, at forskningen blev udført i mangel af kommercielle eller økonomiske forhold, der kunne fortolkes som en potentiel interessekonflikt.
Ashburner, J. Friston, K. J. (2000). Voksel-baseret morfometri-metoderne. Neurobillede 11, 805-821. doi: 10.1006 / nimg.2000.0582
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Bamadis, P. D., Vivas, A. B., Styliadis, C., Klados, M., Schlee, et al. (2014). En gennemgang af fysiske og kognitive indgreb i aldring. Neurosci. Biobehav. Åb 44, 206-220. doi: 10.1016 / j.neubiorev.2014.03.019
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Barulli, D., og Stern, Y. (2013). Effektivitet, kapacitet, kompensation, vedligeholdelse, plasticitet: nye koncepter i cognitice reserve. Tendenser Cogn. Sci. 17, 502–509. doi: 10.1016 / j. tics.2013.08.012
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
sort, J. E., Isaacs, K. R., Anderson, B. J., Alcantara, A. A. og Greenough, V. T. (1990). Læring forårsager synaptogenese, mens motoraktivitet forårsager angiogenese i hjernebarken hos voksne rotter. Proc. Natl. Acad. Sci. USA 87, 5568-5572. doi: 10.1073 / pnas.87.14.5568
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Bliss, T. V. og Lomo, T. (1973). Langvarig potentiering af synaptisk transmission i den dentate af den aneastetiserede kanin efter stimulering af perforeringsbanen. J. Physiol. 232, 331–356. doi: 10.1113 / jphysiol.1973.sp010273
PubMed Abstract / CrossRef fuldtekst / Google Scholar
Boyke, J., Driemeyer, J., Gaser, C., B Prischel, C. og May, A. (2008). Træningsinduceret hjernestruktur ændres hos ældre. J. Neurosci. 28, 7031–7035. doi: 10.1523 / JNEUROSCI.0742-08.2008
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
brun, S., Martins, M. og Parsons, M. (2006). Det neuronale grundlag for menneskelig dans. Cereb. Bark 16, 1157-1167. doi: 10.1093 / cercor / bhj057
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Colcombe, S. J., Erickson, K. I., Scalf, P. E., Kim, J. S., Prakash, R., McAuley, E., Et Al. (2006). Aerob træningstræning øger hjernevolumen hos aldrende mennesker. J. Gerontol. A. Biol. Sci. Middelhavs. Sci. 61, 1166–1170. doi: 10.1093 / gerona / 61.11.1166
PubMed Abstract | CrossRef Fuld tekst/Google Scholar
Ech Privarri, C., Aalten, P., Uyling, H. B. M., Jacobs, H. I. L., Visser, P. J., Gronenschild, M., et al. (2011). Atrofi i parahippocampal gyrus som en tidlig biomarkør for sygdom. Hjerne Struct. Funktion. 215, 265–271. doi: 10.1007 / s00429-010-0283-8
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Edelmann, E., Lessmann, V. og Brigadski, T. (2014). Præ-og postsynaptiske vendinger i BDNF-sekretion og handling i synaptisk plasticitet. Neurofarmakologi 76, 610-627. doi: 10.1016 / j. neuropharm.2013.05.043
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Erickson, K. I., Voss, M. V., Prakash, R. S., Basak, C., Chaddock, L., et al. (2011). Øvelse træning øger størrelsen af hippocampus og forbedrer hukommelsen. Proc. Natl. Acad. Sci. USA 108, 3017-3022. doi: 10.1073 / pnas.1015950108
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Erickson, K. I., Viinstein, A. M., Og L. L. (2012). Fysisk aktivitet, hjernens plasticitet og sygdom. Bue. Middelhavs. Res. 43, 615-621. doi: 10.1016 / j. arcmed.2012.09.008
PubMed Abstract | CrossRef fuldtekst/Google Scholar
FL Larsel, A., Ruschevyh, R., kr larger, K., Vilemer, C., Vinter, B., V Kristilker, K., Et Al. (2010). Fysisk aktivitet og hukommelsesfunktioner: er neurotrofiner og hjernegrå stofvolumen det manglende led? Neuroimage 49, 2756-2763. doi: 10.1016 / j.neuroimage.2009.10.043
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Freret, T., Gaudreau, P., Schumann-Bard, P., Billard, J. M. og Popa-vogner, A. (2015). Mekanismer, der ligger til grund for den neurobeskyttende virkning af hjernereserven mod depression i det sene liv. J. Neural. Transm. (Vienna) 122, S55–S61. doi: 10.1007 / s00702-013-1154-2
PubMed abstrakt / CrossRef Fuld tekst/Google Scholar
Gregory, S. M., Parker, B. og Thompson, P. D. (2012). Fysisk aktivitet, kognitiv funktion og hjernesundhed: hvad er træningstræningens rolle i forebyggelsen af demens? Hjerne Sci. 2, 684–708. doi: 10.3390 / brainsci2040684
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
h Larsnggi, J., Koeneke, S. L., L. og J. L., L. (2010). Strukturel neuroplasticitet i det sensorimotoriske netværk af professionelle kvindelige balletdansere. Hum. Brain Mapp. 31, 1196–1206. doi: 10.1002 / hbm.20928
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Helmstaedter, C., og H. F. (1990). Den verbale Lærings-og fastholdelsestest. Et nyttigt og differentieret værktøj til evaluering af verbal hukommelse ydeevne. – Nej. Bue. Neurol. Psykiatr. (1985) 141, 21–30.
PubMed Abstract/Google Scholar
Huang, E. J. og Reichardt, L. F. (2001). Neurotrophiner: roller i neuronal udvikling og funktion. Annu. Pastor Neurosci. 24, 677–736. doi: 10.1146 / annurev.neuro.24.1.677
PubMed Abstract | CrossRef fuldtekst/Google Scholar
H Larsen, K., Binetti, C., Hamilton, D. A., Stephan, T., Flanagin, V. L., Linn, J., et al. (2011). Strukturel og funktionel plasticitet af hippocampal formation i professionelle dansere og slackliners. Hippocampus 21, 855-865. doi: 10.1002 / hipo.20801
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Karvonen, M. J., Kentala, E., Og Mustala, O. (1957). Virkningerne af træning på puls: en langsgående undersøgelse. Ann. Middelhavs. Eksp. Biol. Fenn. 35, 307–315.
PubMed Abstract / Google Scholar
Kattenstroth, J. C., Kalisch, T., Holt, S., Tegethoff, M. Og Dinse, H. R. (2013). Seks måneders danseintervention forbedrer postural, sensorimotorisk og kognitiv præstation hos ældre uden at påvirke hjerte-respiratoriske funktioner. Front. Aldrende Neurosci. 5:5. doi: 10.3389 / fnagi.2013.00005
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Kattenstroth, J. C., Kolankovska, I., Kalisch, T., og Dinse, H. R. (2010). Overlegen sensorisk, motorisk og kognitiv præstation hos ældre personer med flerårige danseaktiviteter. Front. Aldrende Neurosci. 2:31. doi: 10.3389 / fnagi.2010.00031
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Kempermann, G., Kuhn, H. G. og Gage, F. H. (1997). Flere hippocampale neuroner hos voksne mus, der lever i et beriget miljø. Natur. 386, 493–495. doi: 10.1038/386493a0
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Kirk-Sanches, N. J. og McGough, E. L. (2014). Fysisk træning og kognitiv præstation hos ældre: aktuelle perspektiver. Clin. Interv. Aldring 9, 51-62. doi: 10.2147 / CIA.S39506
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Lessmann, V., og Brigadski, T. (2009). Mekanismer, placeringer og kinetik af synaptisk BDNF-sekretion: en opdatering. Neurosci. Res. 65, 11-22. doi: 10.1016 / j. neures.2009.06.004
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Maass, A., D P., S., Goerke, M., Becke, A., Sobieray, U., Neumann, K., Et Al. (2015). Relationship between peripheral IGF-1, VEGF and BDNF levels and exercise-related changes in memory, hippocampal perfusion and volumes in older adults. Neuroimage 131, 142–154. doi: 10.1016/j.neuroimage.2015.10.084
PubMed Abstract | CrossRef Full Text | Google Scholar
Mattson, M. P. (2008). Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N Y Acad. Sci. 1144, 97–112. doi: 10.1196/annals.1418.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Müller, P., Rehfeld, K., Lüders, A., Schmicker, M., H kurskelmann, A., Kaufmann, J., et al. (2016). Effekter af dans-og sundhedssportstræning på den grå hjernesag hos sunde seniorer. Sportsvæddemål. 46, 213-222. doi: 10.1007 / s12662-016-0411-6
CrossRef Fuld tekst / Google Scholar
Niemann, C., Godde, B. Og Voelcker-Rehage, C. (2014). Ikke kun kardiovaskulær, men også koordineret motion øger hippocampusvolumen hos ældre voksne. Front. Aldrende Neurosci. 6:170. doi: 10.3389 / fnagi.2014.00170
PubMed Abstract / CrossRef fuldtekst/Google Scholar
Osvald, D., Gunselmann, T., Rupprecht, R. og Hagen, B. (2006). Differentielle effekter af enkelt versus kombineret kognitiv og fysisk træning med ældre voksne: SimA-undersøgelsen i et 5-årigt perspektiv. EUR. J. Aldring 3, 179-192. doi: 10.1007 / s10433-006-0035-CrossRef fuldtekst / Google Scholar
Pantel, J., krats, B., Essig, M., Og Schr Krisde, J. (2003). Parahippocampal volumenunderskud hos personer med aldringsassocieret kognitiv tilbagegang. Er. J. Psykiatri 160, 379-382. doi: 10.1176 / appi.ajp.160.2.379
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Park, H., og Poo, M. M. (2013). Neurotrophin regulering af neurale kredsløb udvikling og funktion. Nat. Pastor Neurosci. 14, 7–23. doi: 10.1038/nrn3379
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Rehfeld, K., H Priskelmann, A., Kaufmann, J. og M Kristller, N. G. (2016). “Effekter af en dans vs. en fitnesstræning på hjernens plasticitet, balancepræstation og opmærksomhed hos sunde seniorer: en ny tilgang med SPM 12 til parvis langsgående gruppesammenligning,” i Proceeding-International Conference of Sports and Neuroscience (Magdeburg), 127-138.
sat, P., Cole, M. A., Hardy, D. J. og Rassovsky, Y. (2011). Hjerne og kognitiv reserve: mediator (er) og konstruer validitet, en kritik. J. Clin. Eksp. Neuropsykol. 33, 121–130. doi: 10.1080 / 13803395.2010.493151
PubMed Abstract | CrossRef fuldtekst/Google Scholar
Schega, L., Peter, B., Brigadski, T., Le Larsmann, V., Isermann, B., Hamacher, D., et al. (2016). Effekt af intermitterende normobarisk hypoksi på aerob kapacitet og kognitiv funktion hos ældre mennesker. J. Sci. Middelhavs. Sport 19, 941-945. doi: 10.1016 / j. jsams.2016.02.012
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Taube, M. (2008). Neurofysiologiske tilpasninger som reaktion på balancetræning. Dtsch. Sportmed. 63, 273–277. doi: 10.5960/DSM.2012.030
CrossRef Fuld tekst/Google Scholar
Taubert, M., Draganski, B., Anvandrer, A., M. L., K., Horstmann, A., Villringer, A., et al. (2010). Dynamiske egenskaber ved menneskelig hjernestruktur: læringsrelaterede ændringer i kortikale områder og tilknyttede fiberforbindelser. J. Neurosci. 30, 11670–11677. doi: 10.1523 / JNEUROSCI.2567-10.2010
PubMed abstrakt / CrossRef fuldtekst / Google Scholar
Thomas, A. G., Dennis, A., Bandettini, P. A. og Johansen-Berg, H. (2012). Virkningerne af aerob aktivitet på hjernens struktur. Front. Psychol. 3:86. doi: 10.3389 / fpsyg.2012.00086
PubMed abstrakt | CrossRef Fuld tekst/Google Scholar
Tisserand, D. J., Pruessner, J. C., sans Arigita, E. J., van Bokstel, M. P., Evans, A. C., Jolles, J., et al. (2002). Regionale frontale kortikale volumener falder forskelligt i aldring: en MR-undersøgelse for at sammenligne volumetriske tilgange og vokselbaseret morfometri. Neurobillede 17, 657-669. doi: 10.1016 / s1053-8119 (02)91173-0
PubMed abstrakt | CrossRef fuldtekst | Google Scholar
van Praag, H., Shubert, T., Jao, C. og Gage, F. H. (2005). Motion forbedrer læring og hippocampal neurogenese hos ældre mus. J. Neurosci. 25, 8680–8685. doi: 10.1523 / JNEUROSCI.1731-05.2005
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Verghese, J., Lipton, R. B., kat, M. J., Hall, C. B., Dervy, C. A., Kuslansky, G., et al. (2003). Fritidsaktiviteter og risikoen for demens hos ældre. N. Engl. J. Med. 348, 2508–2516. doi: 10.1056 / NEJMoa022252
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Vital, T. M., Stein, A. M., De Melo Coelho, F. G., Arantes, F. J., Teodorov, E., Santos-Galdur, R. F., et al. (2014). Fysisk træning og vaskulær endotelvækstfaktor (VEGF) hos ældre: en systematisk gennemgang. Bue. Gerontol. Geriatr. 59, 234–239. doi: 10.1016 / j. archger.2014.04.011
PubMed abstrakt | CrossRef fuldtekst/Google Scholar
Voelcker-Rehage, C., og Niemann, C. (2013). Strukturelle og funktionelle hjerneændringer relateret til forskellige typer fysisk aktivitet i hele levetiden. Neurosci. Biobehav. Åb 37, 2268-2295. doi: 10.1016 / j.neubiorev.2013.01.028
PubMed Abstract | CrossRef fuldtekst/Google Scholar
R. J., Fields, R. D. og Johansen-Berg, H. (2012). Plasticitet i grå og hvid: neuroimaging ændringer i hjernens struktur under læring. Nat. Neurosci. 15, 528–536. doi: 10.1038 / nn.3045
PubMed abstrakt / CrossRef Fuld tekst / Google Scholar
Simmermann, P., og Fimm, B. (2002). Test batteri til opmærksomhedstest (TAP), version 1.7, manual—Part1. P. R. S.: Psytest.