
rak płaskonabłonkowy (SCC), znany również jako rak kolczasty, jest jedną z głównych przyczyn zgonów związanych z rakiem na całym świecie. SCC jest złośliwym nowotworem nabłonkowym, powstającym w tkankach, które stanowią barierę między organizmem a środowiskiem, takimi jak skóra, jama ustna, przełyk i płuca . Na wcześniejszym etapie SCC towarzyszy rogowacenie naskórka i powstawanie wrzodów na powierzchni błony śluzowej, a głębsze tkanki są atakowane przez SCCs w późniejszym etapie. W przypadku mikroskopowego wyglądu komórki SCC ułożone są w gniazda, które są otoczone wielokątnymi komórkami z wyraźnymi granicami komórek i jądrami hiperchromatycznymi. SCCs mają dużą tendencję do przerzutów, zwykle przez regionalne węzły chłonne i mogą powodować systematyczne uszkodzenia wielu narządów. Chirurgia, laseroterapia i promieniowanie nadal są najczęściej stosowanym leczeniem SCCs . Unikalne cechy cytomorfologiczne wariantów SCC prowadzą do odrębnego leczenia i wyników .
antygen powierzchniowy komórki CD109 jest glikozylofosfatydyloinozytolem (GPI) związanym z glikoproteiną o masie około 170 kDa i członkiem rodziny makroglobulin A2 (A2M) / C3, C4, C5 białek zawierających tioester . Ludzki gen CD109 znajduje się w chromosomie 6q, stanowiąc około 3,3% całkowitej sekwencji genomowej CD109 z jej 33 eksonami . Izolowany CD109 cDNA zawiera ramkę o odczycie otwartym 4335 bp kodującą 1445 aminokwasów (aa) . Białko CD109 zawiera 21 aa N-końcowy peptyd prowadzący, przypuszczalny region przynęty (aa 651-683), miejsce wiązania tioestru (aa 918-924), reaktywność tioestru definiującą heksapeptyd (Aa1030–1035) i sekwencję sygnału zakotwiczającego C-końcowy konsensus GPI z rozszczepianiem przewidywanym po aminokwasie 1420 (Fig. 1) . N-końcowy peptyd prowadzący zakotwicza białko w błonie wewnętrznej w obrębie peryplazmy i N-końcowy fragment CD109 wydzielany z komórek po rozszczepieniu przez proteazę furinową . Rozszczepienie proteazy w znacznym stopniu nieuporządkowanym regionie przynęty aktywuje CD109, powodując zmianę konformacyjną, która zatrzymuje proteazę w strukturze przypominającej klatkę i ujawnia wysoce reaktywne Wiązanie tioestrowe. Miejsce wiązania tioestru, które po aktywacji może kowalencyjnie łączyć CD109 z resztami lizyny na powierzchni atakującej proteazy. Poza tym Sekwencja sygnału C-końcowego konsensusu GPI-kotwica jest również znana jako domena wiążąca receptor .

The structure of CD109 protein. Białko CD109 o Pełnej długości Zawiera 1445 aminokwasów (aa), w tym 21 aa N-końcowy peptyd sygnałowy, przypuszczalny region przynęty (aa 651-683), miejsce wiązania tioestru (aa 918-924), reaktywność tioestru definiującą heksapeptyd (aa1030-1035) i sekwencję sygnału zakotwiczającego C–końcowy konsensus GPI (aa 1420-1445) z przewidywanym rozszczepieniem po aminokwasie 1420
cd109 został po raz pierwszy zidentyfikowany jako antygen powierzchniowy komórki przez przeciwciało monoklonalne wyhodowane przeciwko prymitywnej linii limfoidalnej/mieloidalnej kg1a . Wcześniejsze badania wykazały, że CD109 ulega ekspresji w podgrupie płodowych i dorosłych komórek jednojądrzastych szpiku kostnego CD34+, aktywowanych limfoblastów T, aktywowanych płytek krwi, komórek śródbłonka , mezenchymalnych komórek macierzystych i kilku ludzkich linii komórek nowotworowych, ale nie ulega ekspresji w spoczynkowych ludzkich komórkach T, płytkach krwi lub leukocytach krwi obwodowej . W szczególności, CD109 został wykryty w SCCs kilku narządów, w tym płuc, przełyku, macicy szyjki macicy, skóry, penisa, jamy ustnej i pęcherzyka żółciowego . CD109 jest składnikiem systemu receptorów transformującego czynnika wzrostu-β1 (TGF-β1) i negatywnie reguluje sygnalizację TGF-β1 . Badania utraty i wzmocnienia funkcji sugerowały, że CD109 może być molekularnym celem rozwoju nowych leków dla SCCs o różnym pochodzeniu tkankowym . W tym przeglądzie przedstawimy podsumowanie aktualnej wiedzy o CD109 i jego związku z SCCs.
ekspresja CD109 w SCCs
w tkankach nowotworowych CD109 wykryto immunohistochemicznie w SCCs, a także w rakach urotelialnych , czerniakach złośliwych , rakach piersi typu podstawno-podobnego , mięśniakomięsaku , mięsakach nabłonkowych i glejakach . W szczególności Shiraki et al. donoszono o CD109-dodatnich okołonaczyniowych komórkach nowotworowych w ludzkich tkankach glejaka niższego stopnia i w modelu mysim rekapitulował glejaka ludzkiego, co sugeruje kluczową rolę CD109 w tej chorobie. Wcześniejsze badania wykazały, że wysoka ekspresja CD109 w SCCs i ograniczona ekspresja w normalnych komórkach płaskonabłonkowych (Tabela 1). Ponadto, CD109 wykazuje wysoką ekspresję w dobrze zróżnicowanych SCCs, a nie w umiarkowanie lub słabo zróżnicowanych SCCs, stąd poziom ekspresji CD109 jest odwrotnie skorelowany ze stopniem guza.
CD109 i SCC płuc
rak płaskonabłonkowy płuc (Lscc) jest jednym z głównych podtypów drobnokomórkowy rak płuc (NSCLC), i stanowi około 20-30% przypadków nsclcs . Ekspresję CD109 zbadano w rakach ludzkich komórek płuc za pomocą ilościowej RT-PCR, która wykazała znamiennie wyższą ekspresję CD109 w rakach płaskonabłonkowych, ale nie w gruczolakorakach, rakach wielkokomórkowych lub rakach drobnokomórkowych . Ponadto, Sato et al. wykorzystano przeciwciało CD109 do analizy ekspresji CD109 w prawidłowych tkankach raka płuc i płuc. W prawidłowych tkankach płuc ekspresja CD109 ograniczała się głównie do komórek podstawnych nabłonka oskrzelowego i oskrzelowego . W tkankach raka płuc silna aktywność immunologiczna była preferencyjnie wykrywana w LSCCs, ale nie w innych typach raków płuc . Wyniki te wskazują, że CD109 jest szczególnie regulowany w LSCCs.
CD109 i SCC przełyku
rak płaskonabłonkowy przełyku (esophagus squamous cell carcinoma, ESCC) stanowią jedną szóstą wszystkich śmiertelności związanych z nowotworami, częściej występującą w krajach azjatyckich . Rokowanie ESCC pozostaje słabe, a 5-letni wskaźnik przeżycia ESCC wynosi około 14-22% . Ostatnie badania sugerują, że zmieniona ekspresja receptorów TGF-β przyczynia się do progresji ESCC, a podwyższona ekspresja białek hamujących sygnalizację TGF-β koreluje ze złym rokowaniem ESCC . Hashimoto et al. zbadano ekspresję CD109 w tkankach ESCC za pomocą ilościowego RT-PCR, a transkrypcję mRNA CD109 zwiększono u około 50% Escc. Później, poziom białka CD109 w ESCC był badany przez immunohistochemię na tkankowych mikromacierzach (TMA). W niezakażonej tkance przełyku ekspresja CD109 była ograniczona w cytozolu stratyfikowanych komórek nabłonka na słabym poziomie. We wszystkich próbkach ESCC z TMAs, nici złośliwych płaskonabłonkowych komórek nabłonkowych wykazywały silne zabarwienie CD109 w cytozolu. W innych typach komórek nie zaobserwowano dodatniego zabarwienia CD109. Ponadto ekspresja CD109 była wyższa w dobrze i umiarkowanie zróżnicowanych Escc niż w słabo zróżnicowanych .
CD109 i SCC szyjki macicy
rak szyjki macicy (macicy szyjki macicy) stanowi około 12% zgonów związanych z rakiem kobiet . SCCs stanowią 80-85% wszystkich nowotworów szyjki macicy . Zhang et al. wykazano, że ekspresja CD109 była znacząco wyższa w SCCs szyjki macicy niż w gruczolakoraku endometrium. Badali również ekspresję CD109 w pięciu liniach komórkowych raka szyjki macicy i obserwowali wysoki poziom ekspresji CD109 w dwóch liniach komórkowych SCC.
CD109 i skórny SCC
skórny rak płaskonabłonkowy (CSCC) jest drugim najczęstszym typem raka skóry bez czerniaka (NMSC) o stale rosnącej częstości występowania . Chociaż CSCC mają ogólnie korzystne rokowanie, nadal jest około 1,5–2% pacjentów umiera z powodu tej choroby . Ekspresja CD109 w CSCCs została zbadana przez immunohistochemię na TMAs . W normalnej skórze CD109 słabo ulegał ekspresji w warstwie podstawnej komórek naskórka, podczas gdy pasma komórek płaskonabłonkowych złośliwych wykazywały silne zabarwienie CD109. Poza tym ekspresja CD109 była odwrotnie skorelowana z klasami CSCC. Podobnie jak Escc, ekspresja CD109 była wyższa w dobrze i umiarkowanie zróżnicowanych CSCC niż w słabo zróżnicowanych . Rak płaskonabłonkowy prącia (PSCC) jest podtypem CSCCs i rzadko był badany . Dong et al. wykazano, że białko CD109 wykazuje dużą ekspresję w złośliwych komórkach płaskonabłonkowych PSCCs w porównaniu z normalnymi tkankami prącia w TMA. Ponadto, wzór ekspresji jest walidowany na świeżych próbkach chirurgicznych PSCC za pomocą immunofluorescencji, qRT-PCR i western blotting, co sugeruje, że CD109 może być biomarkerem PSCC .
CD109 i SCC jamy ustnej
rak płaskonabłonkowy jamy ustnej (OSCC) stanowi 2-3% wszystkich nowotworów na świecie . Utrata mutacji funkcyjnej receptora TGF-β typu II jest częstym zdarzeniem dla SCC jamy ustnej . Ekspresję CD109 w prawidłowych tkankach jamy ustnej i tkankach OSCC u 124 pacjentów zbadano za pomocą zabarwienia immunohistochemicznego. Wysokie poziomy ekspresji CD109 były często wykrywane w SCCs i zmianach przednowotworowych jamy ustnej, ale nie w normalnym nabłonku płaskonabłonkowym . Ponadto poziom ekspresji CD109 był znacząco wyższy w dobrze zróżnicowanych OSC niż w umiarkowanie lub słabo zróżnicowanych OSC, co oznacza, że ekspresja CD109 jest skorelowana ze stopniami różnicowania OSC . Ponadto, linie komórkowe OSCC nadekspresujące CD109 wykazywały przyspieszony wzrost komórek in vitro, co sugeruje, że CD109 bierze udział w progresji Oscc.
CD109 i SCC pęcherzyka żółciowego
rak pęcherzyka żółciowego (GBC) jest najbardziej agresywnym rakiem dróg żółciowych z najkrótszą medianą przeżycia . Rak płaskonabłonkowy pęcherzyka żółciowego (gscc) stanowi 3% nowotworu złośliwego tego narządu . Podtypy tkanek GBCs, w tym adenocarcinoma (AC), squamous cell carcinoma (SCC) i adenosquamous carcinoma (ASC), były badane na TMAs przez immunohistochemiczne zabarwienie przeciwciałem CD109. Barwienie CD109 było ujemne we wszystkich prawidłowych tkankach pęcherzyka żółciowego i tkankach AC. Tymczasem komórki cd109 dodatnie stwierdzono w 86,7% SCCs i 91,7% ASC. Ponieważ CD109 jest wyraźnie wyrażony w złośliwych komórkach sqamous w pęcherzyku żółciowym, CD109 może być markerem diagnostycznym dla pęcherzyka żółciowego SCCs i ASCs .
szlak sygnałowy TGF-β i SCCs
szlak sygnałowy TGF-β jest zaangażowany w wiele procesów komórkowych, w tym wzrost komórek, apoptozę różnicowania komórek i homeostazę komórkową . Rodzina ligandów TGF-β, TGF-β1, TGF-β2 i TGF-β3 wiąże się ze specyficznymi receptorami kinazy serynowo-treoninowej typu I i typu II (TGF-ßR1 i TGF-ßR2) , co powoduje aktywację aktywności kinazy TGF-ßR1 . Aktywowany TGF-ßR1 następnie propaguje sygnał przez fosforylowanie jego wewnątrzkomórkowych substratów, R-SMADs (Smad2 i Smad3). Smad2 i Smad3 wchodzą w interakcje z TGF-ßR1 i SARA (Kotwica Smad do aktywacji receptora), białkiem domeny FYVE, które oddziałuje bezpośrednio z Smad2 i Smad3, SARA działa w celu rekrutacji Smad2 do receptora TGF-β, a następnie fosforylowane R-Smady tworzą heteromeryczne kompleksy z Co-SMAD (Smad4) . Po fosforylacji i kolejnych kompleksach ze Smad4, te kompleksy R-Smads są uwalniane z TGF-ßR1 i SARA, a następnie translokują się do jądra, gdzie wchodzą w interakcje z czynnikami transkrypcyjnymi, które rekrutują je do określonych elementów promotorowych genów docelowych .
endocytoza receptora jest kluczowym mechanizmem regulacyjnym w transdukcji sygnału. Receptory TGF-β są internalizowane zarówno przez drogi zależne od klatryny, jak i caveolae. Internalizacja receptorów TGF-β przez doły pokryte klatryną została połączona z sygnalizacją przez smad2/3 i recyklingiem receptorów. Natomiast lokalizacja receptora TGF-β w caveolae wiąże się z obniżeniem sygnalizacji Smad2/3 i degradacją receptora po ubikwitynacji przez ligazę E3-ubikwityny Smerf2 . Jednak hamujące Smady (Smad6 i Smad7) tworzą odrębną podklasę Smad, które działają w sposób przeciwny do R-Smad i antagonizują sygnalizację . Mogą konkurować z R-Smads o wiązanie z aktywowanym TGF-ßR1, a tym samym hamować fosforylację R-Smads . Ponadto, rekrutują ligazy E3-ubikwityny do aktywowanego TGF-ßR1, powodując ubikwitynację i degradację receptora oraz zakończenie sygnalizacji .
Dysregulacja szlaku TGF-β jest związana z wieloma typami raka . Badania wykazały, że sygnalizacja TGF-β wywołuje efekt zapobiegawczy we wcześniejszych stadiach nowotworu, ale efekt supresyjny podczas późniejszego rozwoju guza . Mutacje w genie TGF-ßR1 stwierdzono również w SCCs skóry, co sugeruje, że inaktywacja TGF-β prowadzi do inicjacji SCCs .
szlak sygnałowy CD109 i TGF-β
CD109 jest Ko-receptorem TGF-β i moduluje aktywność receptora sygnałowego TGF-β w sposób specyficzny dla komórki . Na powierzchni komórki CD109 negatywnie reguluje szlak sygnałowy TGF-β1 poprzez tworzenie kompleksu receptorowego z TGF-ßR1 i TGF-ßR2 w ludzkich keratynocytach . Receptory TGF-β ulegają degradacji po ubikwitynacji przez ligazę E3-ubikwityny Smerf2 i są internalizowane zarówno przez drogi zależne od klatryny, jak i zależne od caveolae . Bizet i in. wykazano, że CD109 wiąże się z caveolin-1 i promuje endocytozę receptora TGF-β. Ponadto CD109 Promuje lokalizację receptorów TGF-β w komorze szczelinowej w obecności ligandu i ułatwia degradację receptora TGF-β. CD109 reguluje również lokalizację i skojarzenie Smad7 / Smerf2 z TGF-ßR1. Hamujące działanie CD109 wymaga ekspresji Smad7 i aktywności ligazy ubikwityny smerf2 . Ponadto CD109 może być uwalniany z powierzchni komórki przez lipazy komórkowe, takie jak fosfatydyloinozytolowa fosfolipaza C (PI-PLC). Rozpuszczalna postać CD109 zachowuje zdolność wiązania TGF-β1 i konfiskuje ją z receptorów TGF-β .
jednak Vorstenbosch et al. stwierdzono, że drogi ALK1-smad1/5 indukowane TGF-β przez CD109 różnią się od szlaków ALK1-Smad1/5 indukowanych przez TGF-β w porównaniu z ALK5-Smad2 / 3 (wszystkie ALK1 i ALK5 są receptorami TGF-β typu i). Odkryli, że sygnalizacja TGF-β hamuje proliferację i migrację komórek śródbłonka, podczas gdy sygnalizacja TGF-β indukuje również te procesy za pomocą ALK1-Smad1/5 . Wykazują one, że ALK1 ulega ekspresji i współlokalizuje się z CD109 w keratynocytach myszy i że myszy nadekspresujące CD109 w naskórku wykazują zwiększoną sygnalizację ALK1-Smad1/5, ale zmniejszoną sygnalizację ALK5-Smad2/3 .
poza tym TGF-β1 jest silnym inhibitorem wzrostu w większości komórek nabłonkowych . Hagiwara et al. wykazano, że doustne linie komórkowe SCC nadekspresja CD109 przyspieszały proliferację komórek i osłabiały działanie antyproliferacyjne pośredniczone przez TGF-β1. W przeciwieństwie do tego, komórki SCC z knockdown CD109 wykazywały wolniejszy wzrost komórek . Wysoki poziom ekspresji CD109 hamował fosforylację Smad2, a tym samym atenuował sygnalizację TGF-β1/Smad2 i upośledzał hamowanie wzrostu komórek za pośrednictwem TGF-β1, knockdown CD109 zwiększał fosforylację Smad2 przez stymulację TGF-β1 . Chociaż CD109 reguluje również sygnalizację Smad1 / 5, nie ma to związku z rozwojem SCC. Łącznie CD109 ułatwia rozwój SCCs poprzez hamowanie szlaku TGF-β-Smad2 / 3 (Fig. 2).

sygnalizacja CD109 i TGF-β. TGF-β wiąże TGF-ßR2, który rekrutuje i fosforyluje TGF-ßR1. Aktywowany TGF-ßR1 propaguje sygnał w dół przez bezpośrednie fosforylowanie Smad2 i Smad3. Fosforylowane R-Smady następnie tworzą heteromeryczne kompleksy z Smad4, w połączeniu z czynnikami transkrypcyjnymi, regulują transkrypcję genów i funkcję komórek. Receptory TGF-β internalizują się poprzez doły pokryte klatryną lub szlak caveolar. CD109 zwiększa Wiązanie TGF-β z receptorami TGF-β i promuje lokalizację receptora TGF-β do caveolae, ostatecznie zwiększa endocytozę receptora TGF-β i ułatwia degradację receptora TGF-β. Hamujące działanie CD109 wymaga ekspresji Smad7 i aktywności ligazy ubikwityny smerf2
sygnalizacja CD109 i STAT3
przetwornik sygnału i aktywator czynnika transkrypcyjnego 3 (STAT3) ma kluczowe znaczenie dla proliferacji, różnicowania, migracji, przeżycia i onkogenezy komórek . Litwinow i in. stwierdzono, że ekspresja białka CD109 była znacznie zmniejszona w łuszczycowym naskórku w porównaniu do sąsiedniej, niezaangażowanej skóry. Jednakże ekspresja mRNA cd109 pozostaje niezmieniona w blaszkach łuszczycowych w porównaniu z normalną skórą, co sugeruje możliwość zwiększenia uwalniania białka CD109 w łuszczycowych keratynocytach . Zasugerowali oni, że wydany/rozpuszczalny CD109 jest w stanie indukować zmiany molekularne, o których wiadomo, że występują w łuszczycy . In vitro odkryli, że transfekcja cd109 siRNA reguluje w dół STAT3, uwalniając CD109 z powierzchni komórek hodowanych ludzkich keratynocytów. Ponadto egzogenny / rekombinowany CD109 indukuje sygnalizację STAT3 w ludzkich keratynocytach . Poza tym, Chuang i in. stwierdzono, że ekspresja CD109 była dramatyczna w komórkach gruczolakoraka płuc z przerzutami, a komórki wykazujące ekspresję cd109 shRNA (shCD109) wykazywały dramatyczne zmniejszenie fosforylacji STAT3. STAT3 knockdown znacznie zmniejszyło przerzuty, a przywrócenie aktywności STAT3 zwiększyło zdolność komórek ekspresji shCD109 do przerzutów . Po aktywacji, STAT3 jest fosforylowany przez nie-receptorowe białkowe kinazy tyrozynowe janus kinase 2 (JAK2), co prowadzi do powstania dimeru STAT3 i translokacji do jądra . Jednakże hamowanie aktywności kinazy JAK u fibroblastów wykazujących nadekspresję CD109 zmniejszyło fosforylowany STAT3 do poziomu podobnego do tego w komórkach rodzicielskich wykazujących niski poziom CD109, co sugeruje, że indukowana przez CD109 fosforylacja STAT3 wymaga aktywności kinazy JAK. Tak więc sygnalizacja JAK / STAT3 może pośredniczyć w wpływie CD109 na wzrost guza i przerzuty .
mimo , że obniżenie poziomu CD109 w ludzkich keratynocytach i komórkach gruczolakoraka płuc obniża poziom sygnalizacji STAT3 in vitro, myszy z niedoborem CD109 wykazywały przeciwne wyniki. Mii i in. wygenerowano myszy z niedoborem CD109, które wykazywały nieprawidłowości skóry, w tym rozrost nabłonka i naciek komórek zapalnych. Donoszono, że fosforylacja STAT3 u myszy z niedoborem CD109 była znacznie wyższa w porównaniu z myszami typu dzikiego. Ponadto podwyższona Regulacja sygnalizacji STAT3 wiąże się ze zwiększoną proliferacją i zaburzonym różnicowaniem keratynocytów .
rozbieżność wyników badań in vitro i In vivo może być spowodowana systematycznymi zmianami mikrośrodowiska w tkankach myszy z niedoborem CD109. Utrata CD109 we wszystkich komórkach myszy może modyfikować podskórne mikrośrodowisko, które aktywuje sygnalizację STAT3 w keratynocytach. Ponadto CD109 może wywierać różne skutki regulacyjne w różnych typach komórek, prowadząc do specyficznej dla danego typu komórki modyfikacji sygnalizacji STAT3. Oprócz keratynocytów CD109 ulega ekspresji w komórkach śródbłonka, nabłonka i fibroblastach, które uczestniczą w budowie tkanki skórnej. Jednak do tej pory związek sygnalizacji CD109 i STAT3 nie został zbadany w tych typach komórek.
CD109 i sygnalizacja EGFR
receptor naskórkowego czynnika wzrostu (EGFR) należy do rodziny receptorów ErbB. Po związaniu ligandem przez EGF, EGFR tworzy dimery, homodimery lub heterodimery z innym członkiem rodziny ErbB HER2 . Dimeryzowane receptory automatycznie fosforylują się nawzajem, a następnie fosforylują białkową kinazę tyrozynową C-Src, która aktywuje STAT3 . Aktywacja EGFR sprzyja migracji komórek, przetrwaniu i proliferacji. W nowotworach złośliwych nadmierna ekspresja EGFR jest skorelowana z głębokością inwazji guza i związana z gorszym rokowaniem . Mutacje prowadzące do nadekspresji EGFR są wykrywane w SCC płuc, SCC głowy i szyi oraz SCC przełyku . CD109 zakotwiczony w błonie w komórkach SK-MG – 1 bezpośrednio oddziałuje z EGFR i wzmacnia sygnalizację EGF, co następnie zwiększa migrację i inwazję komórek, podczas gdy wydzielany CD109 nie ma wpływu na sygnalizację EGF . EGFR może pośredniczyć w wpływie CD109 na sygnalizację STAT3, co wymaga dalszych badań w celu wyjaśnienia (rys. 3).
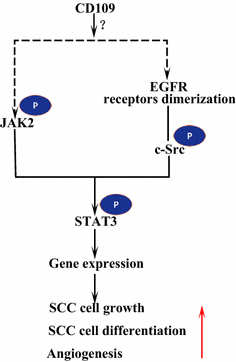
sygnalizacja CD109 i EGFR / STAT3. CD109 ułatwia sygnalizację EGFR. Dimeryzowane receptory EGFR fosforylują C-Src, który z kolei fosforyluje STAT3. CD109 może również aktywować STAT3 poprzez fosforylację JAK2, a następnie promować wzrost komórek SCC, różnicowanie i angiogenezę
do tej pory szeroki obraz nowotworu i rozwoju SCC pozostają niekompletne. Pomiędzy TGF-β a STAT3 lub EGFR występują połączenia krzyżowe. CD109 jest prawdopodobnie jednym z kluczowych efektorów sieci sygnalizacyjnej regulującej SCCs. Nie odnotowano jednak bezpośrednich dowodów na określenie roli CD109 w patogenezie SCCs.
perspektywy
badania z próbek tkanek ludzkich wskazują, że CD109 wykazuje dużą ekspresję w SCCs wielu narządów, szczególnie w dobrze zróżnicowanych złośliwych komórkach płaskonabłonkowych . Wykrywając ekspresję CD109 z immunohistochemią w tkankach ludzkich, CD109 może potencjalnie działać jako biomarker w celu określenia progresji SCC. Obecne badania sugerują, że CD109 ma wysoką ekspresję w dobrze zróżnicowanych SCCs, a jego ekspresja jest niższa w niezróżnicowanych SCCs . Nie jest jednak jasne, czy CD109 jest związany z inwazją naczyniową, przerzutami i rokowaniem po operacji. Dlatego potrzebne są dalsze badania w celu zbadania kliniczno-patologicznego znaczenia CD109 w SCCs w większej próbie.
CD109 jest glikoproteiną związaną z GPI, która umożliwia jej uwolnienie z błony . Rozpuszczalna postać CD109 wpływa również na Wiązanie TGF-β z jego receptorami, a następnie moduluje progresję SCC . Litwinow i in. stwierdzono, że CD109 uwalniany z powierzchni komórki do środowiska pozakomórkowego, a uwolniona forma CD109 zachowuje swoją zdolność do indukowania wewnątrzkomórkowych szlaków sygnałowych. Poza tym, Sakakura i in. poinformował, że serum CD109 zostało uwolnione przez xenografted tumor i zwiększa się proporcjonalnie do objętości xenograft guza. Dlatego wykrywanie poziomu cd109 w surowicy może pomóc w monitorowaniu nadekspresji nowotworów CD109, w tym SCCs.
Exosomy komunikują pierwotną zmianę nowotworową i jej niszę za pośrednictwem pakietu zawierającego wybrane białka lub inne cząsteczki . Izolacja i analizy krążącego egzosomu związanego z nowotworem mogą służyć jako biomarkery do diagnozowania pacjentów z rakiem. Egzogenny CD109 został zidentyfikowany jako składnik egzosomu wydzielanego z transfekowanych 293 komórek, co czyni go obiecującym celem diagnostyki egzosomów . Mimo to, do tej pory nie odnotowano CD109 w egzosomach pochodzących z SCC lub innych komórek nowotworowych. Podstawowe mechanizmy pakowania CD109 do exosomów zasługują na bardziej szczegółowe badanie.
oprócz potencjału jako biomarkera, CD109 może być celem dla podejść terapeutycznych. CD109 jest białkiem błonowym, które może być skierowane bezpośrednio przez specyficzne przeciwciała lub enzymy. CD109 może być również rozpoznawany przez pojazdy docelowe do dostarczania leków. Jednak szczegółowe role CD109 w patogenezie SCCs są nadal niejasne. Na przykład należy zdefiniować zależność niższego wyrażenia CD109 i niezróżnicowanego SCCs. Interwencja CD109 może być rozważana w praktyce klinicznej dopiero po dokładnej ocenie ryzyka i korzyści.