
Plateepitelkarsinom (SCC), også kjent som prickle celle carcinoma, er en av de viktigste årsakene til kreftrelatert død i verden. SCC er en ondartet epitelial tumor, oppstår i vev som gir en barriere mellom en organisme og miljøet, som hud, munn, hulrom, spiserør og lunge . PÅ et tidligere stadium er SCC ledsaget av epidermal keratinisering og sårdannelse i slimhinneoverflaten, og de dypere vevene blir invadert Av SCCs på et senere stadium. For mikroskopisk utseende er SCC-cellene arrangert i reir, som er omgitt av polygonale celler med forskjellige cellegrenser og hyperkromatiske kjerner. SCCs har en høy tendens til å metastasere, vanligvis gjennom regionale lymfeknuter, og kan forårsake systematiske skader på flere organer. Kirurgi, laserterapi og stråling fortsetter å være den mest brukte behandlingen For SCCs . De unike cytomorfologiske egenskapene til scc-varianter fører til distinkt behandling og utfall .CD109 Er et glykosylfosfatidylinositol (GPI)-bundet glykoprotein på ca .170 kDa og medlem av a2-makroglobulin (a2M)/C3, C4, C5-familien av tioesterholdige proteiner. Det humane CD109-genet befinner seg i kromosom 6q, og utgjør omtrent 3,3% AV DEN totale CD109 – genomsekvensen med sine 33 eksoner . Den isolerte CD109 cDNA består av en 4335 bp åpen lesing ramme koding en 1445 aminosyre (aa) . CD109-proteinet inneholder et 21 aa N-terminallederpeptid, en antatt agnregion (aa 651-683), et tioesterbindingssted (aa 918-924), en tioesterreaktivitet som definerer heksapeptid (aa1030–1035) og En C-terminal konsensus GPI-ankersignalsekvens med spaltningen spådd å oppstå etter aminosyre 1420 (Fig. 1) . Det n-terminale lederpeptidet forankrer proteinet til den indre membranen i periplasma og Det N-terminale fragmentet AV CD109 utskilt fra celler etter spaltning av furinproteasen . Proteasespaltningen av en stort sett uordnet agnregion aktiverer CD109, noe som resulterer i en konformasjonsendring som fanger proteasen i en burlignende struktur og eksponerer den svært reaktive tioesterbindingen. Tioesterbindingsstedet som ved aktivering kan kovalent koble CD109 TIL lysinrester på overflaten av den angripende proteasen. Dessuten Er C-terminal konsensus GPI-anker signal sekvens også kjent som reseptor-bindende domene .

The structure of CD109 protein. CD109-proteinets fulle lengde består av 1445 aminosyre (aa), inkludert et 21 aa N-terminalt signalpeptid, en antatt agnregion (aa 651-683), et tioesterbindingssted (aa 918-924), en tioesterreaktivitet som definerer heksapeptid (aa1030–1035) og En C-terminal konsensus GPI-ankersignalsekvens (aa 1420-1445) med spaltningen spådd å oppstå etter aminosyre 1420
cd109 ble først identifisert som et celle-overflateantigen ved et monoklonalt antistoff hevet mot den primitive lymfoide/myeloide cellelinjen kg1a . TIDLIGERE studier rapporterte AT CD109 er uttrykt på en undergruppe AV føtale OG voksne CD34 + positive benmargsmononukleære celler, aktiverte t-lymfoblaster, aktiverte blodplater, endotelceller, mesenkymale stamcelleundergrupper og flere humane tumorcellelinjer, men ikke uttrykt i hvilende humane t-celler, blodplater eller perifere blodleukocytter . SPESIELT HAR CD109 blitt påvist I SCCs av flere organer, inkludert lunge, spiserør, livmorhals, hud , penis, munnhule og galleblæren . CD109 er en del av det transformerende vekstfaktoren-β 1 (TGF-β 1) reseptorsystemet og regulerer NEGATIVT TGF-β 1 signalering . Tap og gevinst av funksjonsstudier antydet AT CD109 kan være et molekylært mål for utvikling av nye terapeutiske Midler for SCCs av forskjellig vevsopprinnelse . I denne gjennomgangen vil vi presentere et sammendrag av DAGENS kunnskap OM CD109 og dets forhold Til SCCs.
CD109-uttrykk I SCCs
I tumorvev ble CD109 immunhistokjemisk påvist i SCCs så vel som uroteliale karsinomer , maligne melanomer , basallignende brystkarsinomer , myxofibrosarkom , epitelsarkomer og gliomer . Spesielt Shiraki et al. RAPPORTERTE CD109-positive perivaskulære tumorceller i humant lavere grad gliomvev og i en musemodell rekapitulert human gliom, noe som tyder PÅ EN nøkkelrolle FOR CD109 for denne sykdommen. Tidligere studier har vist at DET høye uttrykket FOR CD109 i SCCs og det begrensede uttrykket i normale plateepitelceller (Tabell 1). VIDERE ER CD109 sterkt uttrykt i godt differensierte SCCs i stedet for i moderat Eller dårlig differensierte SCCs, og dermed er uttrykksnivået FOR CD109 omvendt korrelert med tumorgrad .
CD109 OG lunge SCC
Lunge plateepitelkarsinom (LSCC) Er en av de viktigste undertyper av ikke-plateepitelkarsinom .småcellet lungekreft (nsclc), og står for ca 20-30% av tilfellene av nsclcs. CD109-ekspresjon ble undersøkt i humane lungecellekarsinomer ved kvantitativ RT-PCR, som viste et signifikant høyere ekspresjon AV CD109 i plateepitelkarsinomer, men ikke i adenokarsinomer, storcellekarsinomer eller småcellekarsinomer . I tillegg Sato et al. BRUKT ET CD109-antistoff for å analysere CD109-uttrykk i normalt lunge-og lungekarsinomvev. I normalt lungevev VAR CD109-ekspresjonen hovedsakelig begrenset til basale celler i bronkial og bronkiolar epitel . I lungekarsinomvev ble sterk immunoreaktivitet fortrinnsvis påvist i LSCCs, men ikke i andre typer lungekarsinomer . DISSE funnene indikerer AT CD109 er spesielt opp-regulert I LSCCs.
CD109 og esophagus SCC
Esophagus squamous cell carcinoma (ESCC) står for en sjettedel av all kreftrelatert dødelighet, som forekommer med høyere forekomst I Asiatiske land . Prognosen FOR ESCC forblir dårlig, og 5-års overlevelse AV ESCC er ca 14-22% . Nyere studier tyder på at endret ekspresjon av tgf-β reseptorer bidrar TIL ESCC progresjon, og forhøyet ekspresjon av hemmende proteiner AV tgf-β signalering korrelerer med dårlig prognose FOR ESCC . Hashimoto et al. undersøkt CD109 uttrykk I ESCC vev ved kvantitativ RT-PCR, OG CD109 mRNA transkripsjon ble oppregulert i ca 50% Av ESCCs. Senere ble proteinnivået AV CD109 i ESCC undersøkt ved immunhistokjemi på vevsmikroarrays (TMA) . I det ikke-syke esophagealvevet var CD109-uttrykket begrenset i cytosol av de stratifiserte epitelceller på et svakt nivå. I ALLE ESCC-prøvene fra Tma-ene viste tråder av ondartede plateepitelceller sterk CD109-farging i cytosolen. INGEN CD109 positiv farging ble observert i andre celletyper. VIDERE VAR CD109 – uttrykket høyere i godt og moderat differensierte ESCCs enn de dårlig differensierte .
CD109 OG livmorhalskreft SCC
livmorhalskreft utgjør ca 12% av kreftrelatert død av kvinner . SCCs står for 80-85% av alle livmorhalskreft . Zhang et al. VISTE AT CD109-uttrykket var signifikant høyere i cervical SCCs enn i endometrial adenokarsinomer. De undersøkte OGSÅ CD109-uttrykk i fem humane livmorhalskreftcellelinjer, og observerte høye NIVÅER AV CD109-uttrykk i to SCC – cellelinjer.
CD109 og kutant SCC
Kutant plateepitelkarsinom (CSCC) er den nest vanligste typen ikke-melanom hudkreft (NMSC) med en stadig økende forekomst . SELV OM CSCC har en generelt gunstig prognose, er det fortsatt ca 1,5–2% av pasientene dør av denne sykdommen . CD109-uttrykk I CSCCs har blitt undersøkt ved immunhistokjemi på TMAs . I de normale skinnene BLE CD109 svakt uttrykt i det basale laget av epidermale celler, mens tråder av ondartede squamous epidermale celler viste sterk CD109-farging. DESSUTEN VAR CD109-uttrykket omvendt korrelert MED CSCC-karakterer. Som ESCCs var uttrykket FOR CD109 høyere i godt og moderat differensierte CSCCs enn de dårlig differensierte . Penile plateepitelkarsinom (PSCC) er en subtype Av CSCCs og har sjelden blitt studert . Dong et al. vist AT CD109 protein er sterkt uttrykt i maligne plateepitelceller Av PSCCs sammenlignet med normalt penisvev på TMAs. Videre er uttrykksmønsteret validert på friske kirurgiske PSCC-prøver ved immunfluorescens, qRT-PCR og western blotting, noe som tyder på AT CD109 kan være en biomarkør for PSCC .
CD109 og oral hulrom SCC
Oral plateepitelkarsinom (OSCC) står for 2-3% av alle kreftformer over hele verden . Tap av funksjonsmutasjon i tgf-β type II-reseptor er en hyppig hendelse for scc i munnhulen . CD109-uttrykk i normalt oralt vev og OSCC-vev fra 124 pasienter ble undersøkt ved immunhistokjemisk farging. Høye NIVÅER AV CD109-ekspresjon ble ofte påvist I SCCs og premaligne lesjoner i munnhulen, men ikke i normale plateepitel . VIDERE var uttrykksnivået FOR CD109 betydelig høyere i godt differensierte OSCCs enn i moderat eller dårlig differensierte OSCCs, noe som innebærer AT CD109-uttrykk er korrelert med Differensieringstrinnene Til OSCCs . I tillegg viste OSCC-cellelinjer SOM overuttrykker CD109 akselerert cellevekst in vitro, noe som innebar AT CD109 involverer I Utviklingen Av OSCCs.
CD109 og galleblæren SCC
Galleblærekreft (GBC) er den mest aggressive av gallekrefttilfellene med kortest median overlevelse . Gallbladder squamous cell carcinoma (GSCC) står for 3% av den ondartede neoplasma i dette organet . Subtyper Av GBCs vev inkludert adenokarsinom (AC), plateepitelkarsinom (SCC) og adenosquamous carcinoma (ASC) ble undersøkt På TMAs ved immunhistokjemisk farging MED ET CD109 antistoff. CD109-fargingen var negativ i alle normale galleblærevev og AC-vev. I mellomtiden BLE CD109 positive celler funnet i 86.7% Av SCCs og 91.7% Av Asc. SIDEN CD109 er tydelig uttrykt i maligne sqamous celler i galleblæren, KAN CD109 være en diagnostisk markør for galleblæren SCCs og ASCs .
TGF – β signalvei og SCCs
TGF-β signalvei er involvert i mange cellulære prosesser, inkludert cellevekst, celledifferensiering apoptose og cellulær homeostase . Familien til FORETAK-β ligander, FORETAK-β1, FORETAK-β2-og FORETAK-β3, binder seg til spesifikke transmembrane type i og type II serin/threonine kinase reseptorer (FORETAK-ßR1 og FORETAK-ßR2) , noe som resulterer i aktivering av FORETAK-ßR1 kinase aktivitet . Den aktiverte TGF-ß 1 forplanter deretter signalet ved å fosforylere dets intracellulære substrater, R-SMADs (Smad2 og Smad3) . Smad2 og Smad3 interagerer MED TGF-ß 1 OG SARA (Smad anker for reseptoraktivering), ET FYVE-domeneprotein som interagerer direkte med Smad2 og Smad3, SARA fungerer for å rekruttere Smad2 TIL tgf-β reseptoren , deretter fosforylerte R-SMADs danner heteromere komplekser Med Co-SMAD (Smad4) . Etter fosforyleringen og påfølgende komplekser Med Smad4 frigjøres disse r-Smads-kompleksene fra TGF – ß 1 OG SARA, og translokeres deretter inn i kjernen hvor de interagerer med transkripsjonsfaktorer som rekrutterer dem til spesifikke promotorelementer av målgener .
Reseptorendocytose er en sentral reguleringsmekanisme i signaltransduksjon. Tgf-β reseptorer internaliseres via både klathrin-og caveolae-avhengige veier. Internalisering av tgf-β reseptorer via klathrin-belagt groper har vært knyttet til signalering Via Smad2 / 3 og reseptor resirkulering. I kontrast er TGF-β reseptorlokalisering i caveolae forbundet med nedregulering Av Smad2 / 3-signalering og reseptornedbrytning etter ubiquitinligase Av E3-ubiquitinligasen Smurf2 . Imidlertid danner inhibitoriske Smads (Smad6 og Smad7) en distinkt underklasse Av Smads som virker motsatt Til R-Smads og motvirker signalering . De kan konkurrere Med R-Smads for binding til aktivert TGF-ß 1 og dermed hemme fosforyleringen Av R-Smads . I tillegg rekrutterer De e3-ubiquitin-ligaser til den aktiverte TGF-ß 1, noe som resulterer i reseptor ubiquitinering og nedbrytning og avslutning av signalering .
Dysregulering AV tgf-β-banen har vært involvert i flere typer kreft . Studier har vist AT TGF-β signalering gir en forebyggende effekt i de tidligere stadiene av tumorigenese, men en undertrykkende effekt under den senere utviklingen av tumor . Mutasjoner I tgf-ß 1-genet er også funnet i SCCs i huden, noe som tyder på at inaktivering av TGF-β fører til oppstart Av SCCs .
CD109 og TGF-β signalvei
CD109 ER EN tgf-β ko-reseptor, og modulerer TGF-β signalreseptor aktivitet på en cellespesifikk måte . PÅ celleoverflaten regulerer CD109 NEGATIVT TGF-β 1-signalveien via dannelse av et reseptorkompleks MED TGF-ß 1 og TGF-ß 2 i humane keratinocytter . Tgf-β reseptorer nedbrytes etter allestedsnærværende via E3-ubiquitinligasen Smurf2, og internaliseres via både klathrin-avhengige og caveolae-avhengige veier . Bizet et al. vist AT CD109 assosieres med caveolin-1 og fremmer tgf-β reseptorendocytose. I TILLEGG fremmer CD109 lokalisering AV tgf-β reseptorene i kaveolarrommet i nærvær av ligand og letter DEGRADERING AV tgf-β reseptorer. CD109 regulerer også lokalisering Og sammenslutning Av Smad7 / Smurf2 MED TGF-ß 1. DE hemmende effektene AV CD109 krever Smad7 uttrykk og smurf2 ubiquitin ligase aktivitet . VIDERE KAN CD109 frigjøres fra celleoverflaten ved cellulære lipaser som fosfatidylinositolspesifikk fosfolipase C (PI-PLC). DEN oppløselige formen PÅ CD109 beholder sin evne til å binde TGF-β1 og konfiskere den bort fra TGF-β reseptorene .
Vorstenbosch et al. RAPPORTERT AT CD109 differensielt regulerte TGF-β-indusert ALK1-Smad1 / 5 versus ALK5-Smad2 / 3 veier (ALK1 og ALK5 er ALLE tgf-β type i-reseptorer). De fant AT TGF-β signalering hemmer endotelcelleproliferasjon og migrasjon, MENS TGF-β signalering også induserer DISSE prosessene via ALK1-Smad1 / 5 . DE viser AT ALK1 uttrykkes og ko-lokaliserer MED CD109 i musekeratinocytter, og at mus overuttrykker CD109 i epidermis viser forbedret ALK1-Smad1 / 5-signalering, men redusert ALK5-Smad2 / 3-signalering .
DESSUTEN ER TGF-β1 en potent veksthemmer i de fleste epitelceller . Hagiwara et al. viste at orale SCC – cellelinjer overuttrykk CD109 akselererte celleproliferasjon og svekket den antiproliferative effekten mediert AV TGF-β 1. I kontrast viste SCC-celler MED CD109 knockdown langsommere cellevekst . Et høyt NIVÅ AV CD109-ekspresjon hemmet Smad2-fosforylering, og dermed svekket tgf-β1/Smad2-signalering og svekker TGF-β1-mediert undertrykkelse av cellevekst, CD109 knockdown økte Smad2-fosforylering ved tgf-β1 stimulering . SELV OM CD109 også regulerer Smad1 / 5-signalering, har den ikke vært knyttet til utviklingen AV SCC. TIL SAMMEN letter CD109 utviklingen Av SCCs via hemming AV tgf-β-Smad2/3-banen (Fig. 2).

CD109 og tgf-β signalering. TGF-β binder TGF-ß 2, som rekrutterer OG fosforylerer TGF-ß 1. Aktivert TGF-ß 1 forplanter signalet nedstrøms ved direkte fosforylering Av Smad2 og Smad3. Fosforylerte R-SMADs danner deretter heteromere komplekser Med Smad4, kombinert med transkripsjonsfaktorer, regulerer gentranskripsjon og cellefunksjon. Tgf-β reseptorer internaliserer via klathrin-belagte groper eller caveolarbanen. CD109 øker tgf-β binding til TGF-β reseptorer og fremmer tgf-β reseptorlokalisering til caveolae, øker til slutt TGF-β reseptorendocytose og letter DEGRADERING AV tgf-β reseptorer. DE hemmende effektene AV CD109 krever Smad7 uttrykk og smurf2 ubiquitin ligase aktivitet
CD109 OG STAT3 signalering
Signaltransduser og aktivator av transkripsjonsfaktor 3 (STAT3) er kritisk for celleproliferasjon, differensiering, migrasjon, overlevelse og onkogenese . Litvinov et al. RAPPORTERT at ekspresjonen AV CD109 protein var markert redusert i psoriasishuden sammenlignet med tilstøtende uinvolvert hud. CD109 mrna-uttrykk er imidlertid uendret i psoriasisplakk sammenlignet med normal hud, noe som tyder på en mulighet FOR AT CD109-proteinfrigivelsen økes i psoriasiskeratinocytter . De foreslo at utgitt / løselig CD109 er i stand til å indusere molekylære endringer som er kjent for å forekomme i psoriasis . In vitro fant de at transfeksjon AV CD109 siRNA nedregulerer STAT3, frigjøring AV CD109 fra celleoverflaten av dyrkede humane keratinocytter. I tillegg induserer eksogen/rekombinant CD109 STAT3-signalering i humane keratinocytter . I tillegg, Chuang et al. RAPPORTERT AT CD109-uttrykk var dramatisk oppregulert i metastatiske lungeadenokarsinomceller, og celler som uttrykte EN CD109 shRNA (shCD109) viste en dramatisk reduksjon I STAT3 fosforylering. STAT3 knockdown reduserte sterkt metastaser, og restaurering AV STAT3-aktivitet økte evnen til shCD109-ekspresjonsceller til å metastasere . VED aktivering fosforyleres STAT3 av ikke-reseptorproteinet tyrosinkinaser janus kinase 2 (JAK2), som fører til DANNELSEN AV STAT3-dimer og translokasjon i kjernen . Hemming AV JAK-kinaseaktivitet i fibroblaster som overuttrykker CD109 reduserte imidlertid fosforylert STAT3 til et nivå tilsvarende det i foreldrecellene som uttrykte lave NIVÅER AV CD109, noe som tyder på AT CD109-indusert STAT3-fosforylering krever JAK-kinaseaktivitet. DERMED kan JAK/STAT3-signalering formidle effektene AV CD109 i tumorvekst og metastase .
selv om knockdown AV CD109 i humane keratinocytter og lungeadenokarsinomceller nedregulerer STAT3-signalering in vitro, VISTE CD109-mangelfulle mus motsatte resultater. Mii et al. GENERERTE CD109-mangelfulle mus, som viste abnormiteter i huden, inkludert epitelhyperplasi og inflammatorisk celleinfiltrasjon. DE rapporterte AT STAT3 fosforylering I CD109-mangelfulle mus var betydelig høyere sammenlignet med villtype mus. I tillegg er oppregulering AV STAT3-signalering forbundet med økt proliferasjon og nedsatt differensiering av keratinocytter .
avviket mellom resultatene fra in vitro-og in vivo-studier kan skyldes systematiske endringer i mikromiljøet i VEV AV CD109-mangelfulle mus. TAP AV CD109 i alle cellene i mus kan endre det subkutane mikromiljøet som aktiverer STAT3-signalering i keratinocytter. I TILLEGG KAN CD109 utøve forskjellige regulatoriske effekter i forskjellige celletyper, noe som fører til celletypespesifikk modifikasjon I STAT3-signalering. I tillegg til keratinocytter uttrykkes CD109 i endotelceller, epitelceller og fibroblaster, som deltar i utgjør hudvevet. Hittil har imidlertid forholdet MELLOM CD109 og STAT3-signalering ikke blitt utforsket i disse celletypene.
CD109 og EGFR-signalering
Epidermal vekstfaktorreseptor (EGFR) er medlem av erbb-familien av reseptorer. Ved ligandbinding av EGF danner EGFR dimerer, enten homodimerer eller heterodimerer med et annet medlem AV erbb-familien HER2 . De dimeriserte reseptorene fosforylerer hverandre og fosforylerer deretter ikke-reseptorproteinet tyrosin c-Src kinase, som aktiverer STAT3 . AKTIVERING AV EGFR fremmer cellemigrasjon, overlevelse og spredning. I maligne tumorer er EGFR overuttrykk korrelert med dybden av invasjon av svulsten og knyttet til dårligere prognose . Mutasjoner som fører TIL EGFR overuttrykk, oppdages i lunge SCC, hode OG nakke SCC og spiserør SCC . Den membranforankrede CD109 I SK-MG-1-celler interagerer direkte MED EGFR og forbedrer EGF-signalering, som senere øker cellemigrasjon og invasjon, mens den utskårne CD109 ikke har noen effekt PÅ EGF-signalering . EGFR kan formidle effekten AV CD109 PÅ STAT3-signalering, noe som krever videre studier for å belyse (Fig. 3).
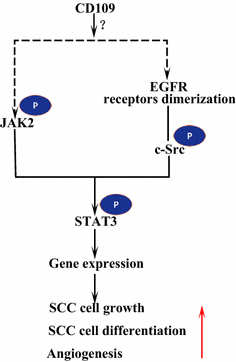
CD109 og EGFR / STAT3 signalering. CD109 forenkler EGFR-signalering. DE dimeriserte EGFR-reseptorene fosforylerer c-Src, som igjen fosforylerer STAT3. CD109 kan også aktivere STAT3 gjennom fosforylering AV JAK2, og deretter fremme SCC cellevekst, differensiering og angiogenese
hittil er det brede bildet av tumorigenese og UTVIKLING AV SCC fortsatt ufullstendig. Det er kryssforhandlinger mellom tgf-β og STAT3 eller EGFR signalveier. CD109 er sannsynligvis en av de viktigste effektorene i signalnettet som regulerer SCCs. Det er imidlertid ikke rapportert direkte bevis for å avgrense ROLLENE TIL CD109 i patogenesen av SCCs.
Perspektiver
Studier fra humane vevsprøver indikerer at CD109 er sterkt uttrykt i SCCs av flere organer, spesielt i godt differensierte maligne plateepitelceller . VED påvisning AV CD109-uttrykk med immunhistokjemi i humant vev, KAN CD109 potensielt virke som en biomarkør for å bestemme progresjonen AV SCC. Nåværende studier tyder på AT CD109 er sterkt uttrykt i godt differensierte SCCs og uttrykket er lavere i udifferensierte SCCs . DET er imidlertid ikke klart om CD109 er assosiert med vaskulær invasjon, metastase og prognose etter operasjon. Derfor er det behov for ytterligere studier for å undersøke DEN kliniske signifikansen AV CD109 i SCCs i en større utvalgsstørrelse.CD109 ER ET gpi-bundet glykoprotein som gjør at DET kan frigjøres fra membranen . DEN oppløselige formen AV CD109 påvirker også bindingen AV TGF-β til dets reseptorer, og modulerer DERETTER scc-progresjon . Litvinov et al. FUNNET AT CD109 frigjort FRA celleoverflaten inn i det ekstracellulære miljøet, og den frigjorte FORM AV CD109 beholder sin evne til å indusere intracellulære signalveier. Dessuten, Sakakura et al. RAPPORTERT at serum CD109 ble utgitt av xenograft tumor og det øker proporsjonalt med volumet av tumor xenograft. Påvisning av serum CD109-nivå kan derfor bidra til å overvåke svulster som overuttrykker CD109, inkludert SCCs.
Eksosomer kommuniserer primær tumorlesjon og nisje via pakningen som inneholder utvalgte proteiner eller andre molekyler . Isolering og analyser av sirkulerende tumorassosiert eksosom kan tjene som biomarkører for diagnostisering av kreftpasienter. Eksogen CD109 har blitt identifisert som en komponent av eksosom utskilt fra transfiserte 293 celler, noe som gjør det til et lovende mål for eksosomdiagnose . LIKEVEL HAR CD109 hittil ikke blitt rapportert i eksosomer avledet FRA SCC eller andre tumorceller. De underliggende mekanismene FOR CD109-emballasje i eksosomer fortjener mer detaljert undersøkelse.I tillegg TIL potensialet som biomarkør, KAN CD109 være et mål for terapeutiske tilnærminger. CD109 er et membranprotein, som kan målrettes direkte av spesifikke antistoffer eller enzymer. CD109 kan også bli gjenkjent av målrettede kjøretøy for levering av legemidler. Imidlertid er de detaljerte rollene TIL CD109 i patogenesen Av SCCs fortsatt uklare. For eksempel må forholdet mellom lavere uttrykk FOR CD109 og utifferentierte SCCs defineres. CD109 intervensjon kan bare vurderes i klinisk praksis etter at risiko og nytte er vurdert nøye.