
扁平上皮癌(SCC)は、プリックル細胞癌としても知られており、世界的にがん関連死の主要な原因の1つです。 SCCは、皮膚、口腔、腔、食道および肺のような生物と環境との間の障壁を提供する組織において生じる悪性上皮腫瘍である。 初期の段階では,SCCは粘膜表面に表皮角化と潰よう形成を伴い,後期にはより深い組織がSccsによって浸潤する。 微視的な外観のために、SCC細胞は巣に配置され、明確な細胞境界および高色素核を有する多角形細胞で囲まれている。 SCCsは、通常、局所リンパ節を介して転移する傾向が高く、複数の臓器の体系的な損傷を引き起こす可能性があります。 外科、レーザー療法および放射はSCCsのための最も広く使用された処置であり続けます。 SCCの変形の独特なcytomorphologic特徴は明瞭な処置および結果をもたらします。
細胞表面抗原CD109は、約170kDaのグリコシルホスファチジルイノシトール(GPI)結合糖タンパク質であり、チオエステル含有タンパク質のa2マクログロブリン(a2m)/C3、C4、C5ファミリーのメンバーである。 ヒトCD109遺伝子は染色体6qに位置し、CD109ゲノム配列全体の約3.3%を33個のエクソンで構成している。 単離されたCD1 0 9cDNAは、1 4 4 5アミノ酸(a a)をコードする4 3 3 5bpの開読フレームを含む。 CD1 0 9タンパク質は、2 1a a N末端リーダーペプチド、推定ベイト領域(a a6 5 1−6 8 3)、チオエステル結合部位(a a9 1 8−9 2 4)、ヘキサペプチド(a a1 0 3 0−1 0 3 5)を定義するチオエステル反応性、およ 1) . N末端リーダーペプチドは、ペリプラズム内の内膜にタンパク質をアンカーし、FURINプロテアーゼによる切断後に細胞から分泌されるCD109のN末端断片。 主に無秩序な餌領域のプロテアーゼの切断は、ケージのような構造にプロテアーゼをトラップし、反応性の高いチオエステル結合を公開するコンフォメーションの変化で、その結果、CD109を活性化します。 活性化に共有結合攻撃プロテアーゼの表面上のリジン残基にCD109をリンクすることができますチオエステル結合部位。 また、C末端コンセンサスGPIアンカーシグナル配列は、受容体結合ドメインとしても知られています。

The structure of CD109 protein. CD109タンパク質の全長は1445アミノ酸(aa)を含み、21aa N末端シグナルペプチド、推定餌領域(aa651-683)、チオエステル結合部位(aa918-924)、ヘキサペプチド(aa1030–1035)とc末端コンセンサスGPIアンカーシグナル配列(aa1420-1445)を定義するチオエステル反応性を含むアミノ酸1420の後に発生すると予測される切断を含む。
cd109は、最初の原始的なリンパ系/骨髄系細胞株kg1aに対して発生したモノクローナル抗体によって細胞表面抗原とし これまでの研究では、cd109は、胎児および成人CD34+陽性骨髄単核細胞、活性化Tリンパ芽球、活性化血小板、内皮細胞、間葉系幹細胞サブセットおよびいくつかのヒト腫瘍細胞株のサブセット上で発現されるが、安静時のヒトT細胞、血小板または末梢血白血球では発現されないことが報告されている。 特に、CD1 0 9は、肺、食道、子宮頸部、皮膚、陰茎、口腔および胆嚢を含むいくつかの器官のSccにおいて検出されている。 CD1 0 9は、形質転換成長因子−β1(TGF−β1)受容体系の構成要素であり、tgf−β1シグナル伝達を負に調節する。 機能の損失と利得の研究は、CD109は、様々な組織起源のSCCsのための新しい治療法の開発のための分子標的である可能性があることを示唆しました。 このレビューでは、CD109の現在の知識とSCCsとの関係の概要を提示します。
SCCsにおけるCD109発現
腫瘍組織では、CD109は、SCCsだけでなく、尿路上皮癌、悪性黒色腫、基底様乳癌、粘液線維肉腫、上皮肉腫および神経膠腫で免疫組織化学 特に、Shiraki e t a l. ヒト低悪性度神経膠腫組織およびマウスモデルで報告されたCD109陽性血管周囲腫瘍細胞は、この疾患のCD109の重要な役割を示唆し、ヒト神経膠腫を 以前の研究では、SccにおけるCD1 0 9の高い発現および正常な扁平上皮細胞における限定的な発現が示されている(表1)。 さらに、CD1 0 9は、中程度または低分化Sccsではなく、高分化Sccsで高度に発現され、したがって、CD1 0 9の発現レベルは、腫瘍の悪性度と反比例する。
CD109および肺SCC
肺扁平上皮癌(LSCC)は、非小細胞肺癌(LSCC)の主要なサブタイNSCLC)、およびnsclcの症例の約20〜30%を占めています。 CD109発現は、扁平上皮癌ではなく、腺癌、大細胞癌または小細胞癌でCD109の有意に高い発現を示した定量的RT-PCRによってヒト肺細胞癌で調べた。 また、Sato e t a l. 正常な肺および肺癌組織におけるCD109発現を分析するためにCD109抗体を使用した。 正常な肺組織では、CD109発現は、気管支および細気管支上皮の基底細胞に主に限定されていた。 肺癌組織では,強い免疫反応性はLsccsで優先的に検出されたが,他のタイプの肺癌では検出されなかった。 これらの知見は、CD109は、特にLSCCsでアップレギュレートされていることを示しています。
CD109および食道SCC
食道扁平上皮癌(ESCC)は、アジア諸国で発生率が高いすべての癌関連死亡率の六分の一を占めています。 ESCCの予後は不良のままであり、ESCCの5年生存率は約14-22%である。 最近の研究は、TGF-β受容体の発現の変化がESCCの進行に寄与し、TGF-βシグナル伝達の阻害タンパク質の発現の上昇はESCCの予後不良と相関することを示唆している。 橋本他 定量的RT-PCRによってESCC組織におけるCD109発現を調べ、CD109mRNA転写は、Esccの約50%でアップレギュレートされました。 その後、ESCCにおけるCD109のタンパク質レベルは、組織マイクロアレイ(TMA)上の免疫組織化学によって調べた。 非罹患食道組織では、CD109発現は弱いレベルで層状上皮細胞の細胞質ゾルに制限されていた。 TMAsからのすべてのESCCサンプルでは、悪性扁平上皮細胞のストランドは、サイトゾルに強いCD109染色を表示しました。 CD109陽性染色は、他の細胞型では観察されなかった。 さらに、CD109発現は、低分化型のものよりもよく、中分化型のESCCsで高かった。
CD109および子宮頸部SCC
子宮頸部(子宮頸部)がんは、女性のがん関連死亡の約12%を占めています。 SCCはすべての子宮頸がんの80-85%を占めています。 張ら CD109発現が子宮内膜腺癌のそれよりも子宮頸部SCCsで有意に高かったことを示した。 彼らはまた、5つのヒト子宮頸癌細胞株におけるCD109発現を調査し、2つのSCC細胞株におけるCD109発現の高レベルを観察した。
CD109および皮膚SCC
皮膚扁平上皮癌(CSCC)は、発生率が絶えず増加している非黒色腫皮膚癌(NMSC)の第二の最も一般的なタイプです。 CSCCは一般的に良好な予後を有するが、患者の約1.5–2%がこの疾患で死亡する。 CsccにおけるCD109発現は、TMAs上の免疫組織化学によって検討されている。 悪性扁平上皮細胞のストランドは強いCD109染色を表示しながら、正常な皮膚では、CD109は、表皮細胞の基底層で弱く発現していた。 また、CD109発現は逆CSCCグレードと相関していた。 ESCCsのように、CD109の発現は、低分化型のものよりもよく、中分化型のCsccで高かった。 陰茎扁平上皮癌(PSCC)はCsccのサブタイプであり、ほとんど研究されていない。 Dongら。 CD109タンパク質は、tma上の正常な陰茎組織と比較してPsccの悪性扁平上皮細胞で高度に発現されることを示した。 さらに、発現パターンは、CD109はPSCCのバイオマーカーである可能性があることを示唆し、免疫蛍光、qRT-PCRとウェスタンブロッティングによって新鮮な外科PSCCサン
CD109および口腔SCC
口腔扁平上皮癌(OSCC)は、世界中のすべての癌の2-3%を占めています。 TGF-β II型受容体における機能変異の喪失は口腔SCCのための頻繁な事象である。 正常な口腔組織および124人の患者からOSCC組織におけるCD109発現は、免疫組織化学染色によって調べた。 高レベルのCD109発現は、SCCsおよび口腔の前悪性病変で頻繁に検出されたが、正常な扁平上皮では検出されなかった。 また、CD109の発現レベルは、CD109発現がOSCCsの分化段階と相関していることを意味する中程度または低分化OSCCsよりもよく分化OSCCsで有意に高かった。 さらに、CD109を過剰発現OSCC細胞株は、CD109はOSCCsの進行に関与することを意味し、in vitroで加速された細胞増殖を示した。
CD109および胆嚢SCC
胆嚢癌(GBC)は、生存期間の中央値が最も短い胆道癌の中で最も攻撃的である。 胆嚢扁平上皮癌(GSCC)は、この器官の悪性新生物の3%を占める。 腺癌(AC)、扁平上皮癌(SCC)、および腺扁平上皮癌(ASC)を含むGBCs組織のサブタイプは、CD109抗体による免疫組織化学染色によってTMAsについて調べた。 CD109染色は、すべての正常な胆嚢組織およびAC組織で陰性であった。 一方、CD109陽性細胞は、SCCsの86.7%およびASCsの91.7%で発見された。 CD109は胆嚢の悪性sqamous細胞ではっきりと発現されるので、CD109は胆嚢SCCsおよびASCsの診断マーカーである可能性があります。
TGF-βシグナル伝達経路とSCCs
TGF-βシグナル伝達経路は、細胞増殖、細胞分化アポトーシス、および細胞恒常性を含む多くの細胞プロセスに関与している。 TGF−βリガンドのファミリー、TGF−β1、TGF−β2およびTGF−β3は、特定の膜貫通i型およびII型セリン/スレオニンキナーゼ受容体(TGF−β R1およびTGF−β R2)に結合し、TGF−β r1キ 活性化されたTGF−β R1は、次いで、その細胞内基質R−Smads(Smad2およびSmad3)をリン酸化することによってシグナルを伝播する。 Smad2とSmad3はTGF-β r1とSARA(受容体活性化のためのSmadアンカー)、Smad2とSmad3と直接相互作用するFYVEドメインタンパク質と相互作用し、SARAはSmad2をTGF-β受容体にリクルートする機能を果たし、リン酸化されたR-SmadはCo-SMAD(Smad4)とヘテロマー複合体を形成する。 Smad4とのリン酸化およびそれに続く複合体の後で、これらのR-Smadsの複合体はtgf-γ r1およびSARAから解放され、そして核に移動しますそれらが標的遺伝子
受容体エンドサイトーシスは、シグナル伝達における極めて重要な調節機構である。 TGF-β受容体は、クラスリン依存性経路とカベオラエ依存性経路の両方を介して内在化される。 クラスリン被覆ピットを介してTGF-β受容体の内在化は、Smad2/3と受容体リサイクルを介してシグナル伝達とリンクされています。 対照的に、カベオラエにおけるTGF-β受容体の局在は、E3-ユビキチンリガーゼSmurf2によるユビキチン化後のSmad2/3シグナル伝達と受容体分解のダウンレ しかし、阻害性Smads(Smad6とSmad7)は、r-Smadsに反対の方法で作用し、シグナル伝達に拮抗するSmadsの別個のサブクラスを形成する。 それらは、活性化TGF−β R1への結合に関してR−Smadsと競合し、したがってR−Smadsのリン酸化を阻害することができる。 さらに、それらは受容器のユビキチン化および低下およびシグナリングの終了に終って活動化させたTGF-γ r1にE3ユビキチンのリガーゼを、募集する。
TGF-β経路の調節不全は、複数のタイプの癌に関与している。 研究は、TGF-βシグナル伝達は、腫瘍形成の初期段階の間に予防効果を誘発するが、腫瘍発生の後期の間に抑制効果を誘発することを示している。 TGF-β r1遺伝子の変異は皮膚のSccにおいても見出されており、TGF-βの不活性化がSccの開始につながることを示唆している。
CD109およびTGF-βシグナル伝達経路
CD109はTGF-β共受容体であり、細胞特異的な方法でTGF-βシグナル伝達受容体活性を調節する。
CD109はtgf-β共受容体であり、細胞特異的な方法でtgf-βシグナル伝達受容体活性を調節する。
細胞表面では、CD109は、ヒトケラチノサイトにおけるTGF-β r1およびTGF-β r2との受容体複合体の形成を介してtgf-β1シグナル伝達経路を負に調節する。 TGF-β受容体は、E3-ユビキチンリガーゼSmurf2によるユビキチン化後に分解し、クラスリン依存性およびカベオラエ依存性経路の両方を介して内在化される。 ビゼー他 CD109がカベオリン-1と結合し、TGF-β受容体エンドサイトーシスを促進することを示した。 さらに、CD109は、リガンドの存在下でcaveolarコンパートメントにTGF-β受容体の局在化を促進し、TGF-β受容体の分解を促進する。 CD109はまた、局在化とSmad7/Smurf2TGF-β r1との関連を調節します。 CD109の阻害効果は、Smad7発現とSmurf2ユビキチンリガーゼ活性を必要とします。 さらに、CD109は、ホスファチジルイノシトール特異的ホスホリパーゼC(PI-PLC)などの細胞リパーゼによって細胞表面から放出することができる。 CD1 0 9の可溶性形態は、TGF−β1に結合し、TGF−β受容体からそれを没収するその能力を保持する。しかし、Vorstenbosch et al. CD1 0 9は、ALK5−Smad2/3経路(ALK1およびALK5は全てtgf−β i型受容体である)に対して、tgf−β誘導性ALK1−Smad1/5を特異的に調節することを報告した。 その結果、tgf-βシグナルは内皮細胞の増殖と遊走を阻害する一方、TGF-βシグナルはALK1-Smad1/5を介してこれらのプロセスを誘導することが分かった。 彼らはALK1が発現し、マウスのケラチノサイトでCD109と共局在し、表皮ディスプレイでCD109を過剰発現するマウスはALK1-Smad1/5シグナル伝達を強化したが、ALK5-Smad2/3シグナル伝達を減少させたことを示している。さらに、TGF-β1は、ほとんどの上皮細胞における増殖の強力な阻害剤である。
さらに、TGF-β1は、ほとんどの上皮細胞における増殖の強力な阻害剤 萩原他 経口SCC細胞株過剰発現CD109は、細胞増殖を加速し、TGF-β1によって媒介される抗増殖効果を障害することを示した。 対照的に、CD109ノックダウンとSCC細胞は遅い細胞増殖を示した。 CD109発現の高レベルは、Smad2のリン酸化を阻害し、このようにtgf-β1/Smad2シグナル伝達を減衰させ、細胞増殖のTGF-β1媒介抑制を損なう、CD109ノックダウンは、TGF-β1刺激によってSmad2のリン酸化を増加させた。 CD109はSmad1/5シグナル伝達も調節しているが、SCCの発達とは関連していない。 一緒に、CD1 0 9は、TGF−β−Smad2/3経路の阻害を介してSccの発生を促進する(図3)。 2).

CD109およびTGF-βシグナル伝達。 TGF−βはTGF−β R2に結合し、これはtgf−β R1を再募集し、リン酸化する。 活性化されたTGF-β r1は、Smad2とSmad3を直接リン酸化することによって下流にシグナルを伝播する。 リン酸化されたR-Smadは、転写因子と組み合わせたSmad4とヘテロマー複合体を形成し、遺伝子転写および細胞機能を調節する。 TGF-β受容体は、クラスリン被覆ピットまたはカベオラル経路を介して内在化する。 CD1 0 9は、TGF−β受容体へのtgf−β結合を増加させ、カベオラエへのTGF−β受容体の局在化を促進し、最終的にTGF−β受容体エンドサイトーシスを増加させ、TGF−β受容体の分解を促進する。 CD109の阻害効果は、Smad7発現とSmurf2ユビキチンリガーゼ活性を必要とします
CD109とSTAT3シグナル伝達
転写因子3(STAT3)のシグナルトランスデューサと活性化因子は、細胞の増殖、分化、遊走、生存、および発癌に重要です。 リトヴィノフ他 CD109タンパク質の発現が著しく隣接する関与していない皮膚と比較して乾癬表皮で減少したことを報告した。 しかし、CD109mRNA発現は、CD109タンパク質放出が乾癬ケラチノサイトで強化されている可能性を示唆し、正常な皮膚と比較して乾癬プラークで変化しません。 彼らは、放出された/可溶性CD109が乾癬で起こることが知られている分子変化を誘導することができることを示唆した。 In vitroでは、CD109siRNAのトランスフェクションがSTAT3、培養ヒトケラチノサイトの細胞表面からのCD109の放出をダウンレギュレートすることがわかった。 さらに、外因性/組換えCD109は、ヒトケラチノサイトにおけるSTAT3シグナル伝達を誘導する。 また、Chuang et al. 転移性肺腺癌細胞においてcd109発現が劇的に上方制御され、CD109shRNA(shcd109)を発現する細胞はSTAT3リン酸化の劇的な減少を示したことを報告した。 STAT3ノックダウンが大幅に転移を減少させ、STAT3活性の回復は、転移するshcd109発現細胞の能力を増加させた。 活性化すると、STAT3は非受容体タンパク質チロシンキナーゼjanusキナーゼ2(JAK2)によってリン酸化され、stat3二量体の形成と核への転座につながる。 しかし、cd109を過剰発現する線維芽細胞におけるJAKキナーゼ活性の阻害は、CD109誘導STAT3リン酸化がJAKキナーゼ活性を必要とすることを示唆し、CD109の低レベ したがって、JAK/STAT3シグナル伝達は、腫瘍増殖および転移におけるCD109の効果を媒介する可能性がある。
ヒトケラチノサイトおよび肺腺癌細胞におけるCD109のノックダウンは、IN vitroでSTAT3シグナル伝達をdownregulatesが、CD109欠損マウスは反対の結果を示した。 Miiら。 上皮過形成、および炎症細胞浸潤を含む皮膚異常を示したCD109欠損マウスを生成しました。 彼らは、CD109欠損マウスにおけるSTAT3のリン酸化が野生型マウスと比較して有意に高かったことを報告した。 さらに、STAT3シグナル伝達のアップレギュレーションは、増加した増殖とケラチノサイトの分化障害に関連付けられています。
IN vitroおよびin vivo研究の結果の不一致は、CD109欠損マウスの組織における微小環境の体系的な変化によって引き起こされる可能性がある。 マウスのすべての細胞におけるCD109の損失は、ケラチノサイトにおけるSTAT3シグナル伝達を活性化する皮下微小環境を変更する可能性があります。 さらに、CD109はSTAT3シグナル伝達における細胞型特異的修飾につながる、異なる細胞型で明確な調節効果を発揮することができます。 ケラチノサイトに加えて、CD109は、皮膚組織を構成する内皮細胞、上皮細胞、および線維芽細胞において発現される。 しかしながら、これまでのところ、CD1 0 9とSTAT3シグナル伝達の関係は、これらの細胞型においては探求されていない。
Cd109およびEGFRシグナル伝達
上皮成長因子受容体(EGFR)は、受容体のErbBファミリーのメンバーである。 EGFによるリガンド結合時に、EGFRは、ErbBファミリー HER2の別のメンバーと二量体、ホモ二量体またはヘテロ二量体のいずれかを形成する。 二量化された受容体は互いに自己リン酸化し、次にSTAT3を活動化させる非受容体蛋白質のチロシンc-Srcのキナーゼをリン酸化します。 EGFRの活性化は、細胞遊走、生存および増殖を促進する。 悪性腫瘍では、EGFRの過剰発現は腫瘍の深達度と相関し、予後不良に関連している。 EGFRの過剰発現につながる突然変異は、肺SCC、頭頸部SCC、および食道SCCにおいて検出される。 SK-MG-1細胞の膜アンカー CD109は直接EGFRと相互作用し、分泌されたCD109はEGFシグナル伝達に影響を与えないが、その後、細胞の遊走と侵入を増加させるEGFシグ EGFRは、STAT3シグナル伝達に対するCD1 0 9の効果を媒介する可能性があり、これを解明するためにさらなる研究を必要とする(図1 4A)。 3).
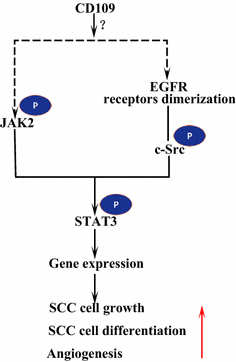
CD109およびEGFR/STAT3シグナリング。 CD109はEGFRシグナル伝達を容易にする。 二量化されたEGFR受容体はc-Srcをリン酸化し、これはSTAT3をリン酸化する。 CD109はまた、JAK2のリン酸化を介してSTAT3を活性化し、SCC細胞の成長、分化、および血管新生を促進することができます
これまでの腫瘍形成およびSCCの発達の広範な画像は不完全なままである。 TGF-βとSTAT3またはEGFRシグナル伝達経路との間には交差トークがある。 CD109は、SCCsを規制するシグナリングネットワークの重要なエフェクターの一つである可能性が高い。 しかし、SCCsの病因におけるCD109の役割を描写する直接的な証拠は報告されていない。
Perspectives
ヒト組織サンプルからの研究は、CD109が多臓器のScc、特に分化した悪性扁平上皮細胞において高度に発現することを示している。
ヒト組織サンプルからの研究は、CD109が多臓器のSccにおいて高度に発現することを示している。 ヒト組織における免疫組織化学によるCD109発現の検出により、CD109は、SCCの進行を決定するためのバイオマーカーとして潜在的に作用する可能性がある。 現在の研究では、CD109は高分化Sccで発現され、その発現は非分化Sccでは低いことが示唆されている。 しかし、CD109が血管浸潤、転移、および手術後の予後と関連しているかどうかは明らかではない。 したがって、より大きなサンプルサイズのSCCsにおけるCD109の臨床病理学的意義を探求するためには、さらなる研究が必要である。CD109はGPI結合糖タンパク質であり、膜から放出することを可能にする。
CD109はGPI結合糖タンパク質であり、膜から放出することを可能にする。
CD109は CD109の可溶性形態はまた、その受容体へのTGF-βの結合に影響を与え、続いてSCCの進行を調節する。 リトヴィノフ他 CD1 0 9が細胞表面から細胞外環境に放出され、放出された形態のCD1 0 9が細胞内シグナル伝達経路を誘導するその能力を保持することを見出した。 また、坂倉らは、””の中で、” 血清CD109は、異種移植腫瘍によって放出され、それは腫瘍異種移植の体積に比例して増加することを報告した。 したがって、血清CD109レベルの検出は、SCCsを含むCD109を過剰発現する腫瘍を監視するのに役立つかもしれない。
エキソソームは、選択されたタンパク質または他の分子を含むそのパッケージを介して原発腫瘍病変およびそのニッチを通信する。 循環腫瘍関連エクソソームの単離および分析は、癌患者の診断のためのバイオマーカーとして役立つ可能性がある。 外因性CD109はそれにexosomeの診断のための有望なターゲットをするトランスフェクトされた293細胞から分泌されるexosomeの部品として識別されました。 さらに、今日まで、CD1 0 9は、SCCまたは他の腫瘍細胞に由来するエキソソームにおいて報告されていない。 エキソソームへのCD109パッケージングの基礎となるメカニズムは、より詳細な調査に値する。
バイオマーカーとしての可能性に加えて、CD109は治療アプローチの標的となる可能性があります。 CD1 0 9は膜タンパク質であり、特異的な抗体または酵素によって直接標的化することができる。 CD109はまた薬剤配達のための目標とされた車によって確認されるかもしれません。 しかし、SCCsの病因におけるCD109の詳細な役割はまだ不明である。 例えば、CD1 0 9の低い発現と未分化Sccとの関係を定義する必要がある。 CD109介在は危険および利点が注意深く評価された後臨床練習でだけ考慮されるかもしれません。