
Il carcinoma a cellule squamose (SCC), noto anche come carcinoma a cellule prickle, è una delle principali cause di morte correlata al cancro in tutto il mondo. SCC è un tumore epiteliale maligno, che si manifesta nei tessuti che forniscono una barriera tra un organismo e l’ambiente, come la pelle, la bocca, la cavità, l’esofago e il polmone . In una fase precedente, SCC è accompagnato da cheratinizzazione epidermica e formazione di ulcera nella superficie della mucosa, e i tessuti più profondi sono invasi da SCC in una fase successiva. Per aspetto microscopico, le cellule SCC disposte in nidi, che sono circondati da cellule poligonali con bordi cellulari distinti e nuclei ipercromatici. I SCC hanno un’alta tendenza a metastatizzare, di solito attraverso i linfonodi regionali, e potrebbero causare danni sistematici a più organi. La chirurgia, la terapia laser e la radiazione continuano ad essere il trattamento più ampiamente usato per SCCs . Le caratteristiche citomorfologiche uniche delle varianti SCC portano a trattamenti e risultati distinti .
L’antigene di superficie cellulare CD109 è una glicoproteina legata al glicosilfosfatidilinositolo (GPI) di circa 170 kDa e un membro della famiglia di proteine contenenti tioesteri a2 macroglobulina (a2M)/C3, C4, C5 . Il gene umano CD109 si trova nel cromosoma 6q, che costituisce circa il 3,3% della sequenza genomica totale CD109 con i suoi 33 esoni . Il cDNA CD109 isolato comprende un frame a lettura aperta da 4335 bp che codifica un aminoacido 1445 (aa). La proteina CD109 contiene un peptide leader N-terminale 21 aa, una regione di esca putativa (aa 651-683), un sito di legame tioestere (aa 918-924), una reattività tioestere che definisce l’esapeptide (aa1030–1035) e una sequenza di segnale di consenso GPI-ancoraggio C-terminale con la scissione prevista dopo l’amminoacido 1420 (Fig. 1) . Il peptide leader N-terminale ancorare la proteina alla membrana interna all’interno del periplasma e il frammento N-terminale di CD109 secreto dalle cellule dopo la scissione dalla proteasi furina . La scissione della proteasi di una regione esca in gran parte disordinata attiva CD109, con conseguente cambiamento conformazionale che intrappola la proteasi in una struttura a gabbia ed espone il legame tioestere altamente reattivo. Il sito di legame tioestere che all’attivazione può collegare covalentemente CD109 ai residui di lisina sulla superficie della proteasi attaccante. Inoltre, la sequenza del segnale di consenso GPI-ancoraggio C-terminale è anche conosciuta come dominio di legame del recettore .

The structure of CD109 protein. Il full-length di CD109 proteina comprende 1445 amminoacido (aa), tra cui un 21 aa N-terminale del peptide segnale, un putativo esca regione (aa 651-683), un sito di legame tioestere (aa 918-924), un tioestere reattività definizione di esapeptide (aa1030–1035) e C-terminale consenso GPI-ancora sequenza segnale (aa 1420-1445) con la scissione previsto a verificarsi dopo aminoacidi 1420
CD109 è stato identificato come una cella dell’antigene di superficie da parte di un anticorpo monoclonale sollevato contro la primitiva linfoidi/cellule mieloidi linea KG1a . Studi precedenti hanno riferito che CD109 è espresso su un sottoinsieme di cellule mononucleate del midollo osseo positivo CD34+ fetale e adulto, linfoblasti T attivati, piastrine attivate, cellule endoteliali , sottoinsiemi di cellule staminali mesenchimali e diverse linee cellulari tumorali umane, ma non espresse in cellule T umane a riposo, piastrine o leucociti del sangue periferico . In particolare, CD109 è stato rilevato in SCC di diversi organi , tra cui polmone , esofago , cervice uterina , pelle , pene, cavità orale e cistifellea . CD109 è un componente del sistema trasformante del ricevitore di fattore di crescita-β1 (TGF-β1) e negativamente regola la segnalazione di TGF-β1 . Studi di perdita e guadagno di funzione hanno suggerito che CD109 può essere un bersaglio molecolare per lo sviluppo di nuove terapie per SCC di varia origine tissutale . In questa recensione, presenteremo una sintesi delle attuali conoscenze di CD109 e del suo rapporto con SCCs.
CD109 espressione in SCCs
Nei tessuti tumorali, CD109 è stato immunoistochimicamente rilevato in SCCs così come carcinomi uroteliali , melanomi maligni , carcinoma mammario basali , mixofibrosarcoma , sarcomi epiteliali e glioma . In particolare, Shiraki et al. le cellule tumorali perivascolari CD109-positive riportate nei tessuti umani di glioma di grado inferiore e in un modello murino hanno ricapitolato glioma umano, suggerendo un ruolo chiave di CD109 per questa malattia. Studi precedenti hanno dimostrato che l’alta espressione di CD109 nel SCC e l’espressione limitata nelle cellule squamose normali (Tabella 1) . Inoltre, CD109 è altamente espresso in SCC ben differenziati piuttosto che in SCC moderatamente o scarsamente differenziati, quindi il livello di espressione di CD109 è inversamente correlato con il grado tumorale .
CD109 polmone e SCC
Polmone carcinoma a cellule squamose (LSCC) è uno dei principali sottotipi di non a piccole cellule del polmone (NSCLC), e rappresenta circa il 20-30% di casi di NSCLCs . L’espressione di CD109 è stata esaminata nei carcinomi a cellule polmonari umane mediante RT-PCR quantitativa, che ha mostrato un’espressione significativamente più elevata di CD109 nei carcinomi a cellule squamose, ma non negli adenocarcinomi, nei carcinomi a grandi cellule o nei carcinomi a piccole cellule . Inoltre, Sato et al. usato un anticorpo CD109 per analizzare l’espressione di CD109 nei tessuti normali del carcinoma del polmone e del polmone. Nei tessuti polmonari normali, l’espressione di CD109 era limitata principalmente alle cellule basali degli epiteli bronchiali e bronchiolari . Nei tessuti del carcinoma polmonare, una forte immunoreattività è stata rilevata preferenzialmente negli LSCC ma non in altri tipi di carcinomi polmonari . Questi risultati indicano che CD109 è specificamente regolato in LSCC.
CD109 e SCC dell’esofago
Il carcinoma a cellule squamose dell’esofago (ESCC) rappresenta un sesto di tutta la mortalità correlata al cancro, che si verifica con una maggiore incidenza nei paesi asiatici . La prognosi di ESCC rimane scarsa e il tasso di sopravvivenza a 5 anni di ESCC è di circa il 14-22%. Studi recenti suggeriscono che l’espressione alterata dei recettori TGF-β contribuisce alla progressione dell’ESCC e l’elevata espressione delle proteine inibitorie della segnalazione TGF-β è correlata alla prognosi infausta dell’ESCC . Hashimoto et al. esaminata l’espressione del CD109 nei tessuti dell’ESCC mediante RT-PCR quantitativa, la trascrizione dell’mRNA CD109 è stata sovraregolata in circa il 50% dei CESC. Successivamente, il livello proteico di CD109 in ESCC è stato esaminato mediante immunoistochimica su microarray tissutali (TMA) . Nel tessuto esofageo non malato, l’espressione CD109 è stata limitata nel citosol delle cellule epiteliali stratificate a un livello debole. In tutti i campioni di ESCC dalle TMA, filamenti di cellule epiteliali squamose maligne hanno mostrato una forte colorazione CD109 nel citosol. Non è stata osservata alcuna colorazione positiva al CD109 in altri tipi di cellule. Inoltre, l’espressione di CD109 era più elevata nei CESC ben e moderatamente differenziati rispetto a quelli scarsamente differenziati .
CD109 e cervice uterina SCC
Il cancro cervicale (cervice uterina) rappresenta circa il 12% della morte correlata al cancro delle donne . Gli SCC rappresentano l ‘ 80-85% di tutti i tumori cervicali . Zhang et al. ha mostrato che l’espressione di CD109 era significativamente più alta nei SCC cervicali rispetto a quella negli adenocarcinomi endometriali. Hanno anche studiato l’espressione di CD109 in cinque linee cellulari di carcinoma cervicale umano e hanno osservato alti livelli di espressione di CD109 in due linee cellulari SCC.
CD109 e SCC cutaneo
Il carcinoma cutaneo a cellule squamose (CSCC) è il secondo tipo più comune di tumore della pelle non melanoma (NMSC) con un’incidenza in costante aumento . Sebbene CSCC abbia una prognosi generalmente favorevole, c’è ancora circa l ‘ 1,5–2% dei pazienti che muoiono per questa malattia . L’espressione di CD109 in CSCCs è stata esaminata dall’immunoistochimica su TMA . Nelle pelli normali, CD109 era debolmente espresso nello strato basale delle cellule epidermiche, mentre i filamenti di cellule epidermiche squamose maligne mostravano una forte colorazione CD109. Inoltre, l’espressione CD109 era inversamente correlata con i gradi CSCC. Come i CSCC, l’espressione di CD109 era più alta nei CSCC ben e moderatamente differenziati rispetto a quelli scarsamente differenziati . Il carcinoma a cellule squamose del pene (PSCC) è un sottotipo di CSCC e raramente è stato studiato . Dong et al. dimostrato che la proteina CD109 è altamente espressa nelle cellule squamose maligne di PSCC rispetto ai normali tessuti del pene su TMA. Inoltre, il modello di espressione è convalidato su campioni PSCC chirurgici freschi mediante immunofluorescenza, qRT-PCR e western blotting, suggerendo che CD109 potrebbe essere un biomarcatore per PSCC .
CD109 e cavità orale SCC
Il carcinoma a cellule squamose orali (OSCC) rappresenta il 2-3% di tutti i tumori in tutto il mondo . La perdita della mutazione della funzione nel recettore TGF-β di tipo II è un evento frequente per la cavità orale SCC . L’espressione di CD109 nei tessuti orali normali e nei tessuti OSC di 124 pazienti è stata esaminata mediante colorazione immunoistochimica. Alti livelli di espressione di CD109 sono stati frequentemente rilevati in SCC e lesioni premaligne del cavo orale, ma non in normali epiteli squamosi . Inoltre, il livello di espressione di CD109 era significativamente più alto negli OSC ben differenziati rispetto agli OSC moderatamente o scarsamente differenziati, il che implica che l’espressione di CD109 è correlata con gli stadi di differenziazione degli OSC . Inoltre, le linee cellulari OSCC che sovraesprimevano CD109 hanno mostrato una crescita cellulare accelerata in vitro , implicando che CD109 coinvolge nella progressione degli OSCC.
CD109 e cistifellea SCC
Il cancro della cistifellea (GBC) è il più aggressivo dei tumori biliari con la sopravvivenza mediana più breve . Il carcinoma a cellule squamose della cistifellea (GSCC) rappresenta il 3% della neoplasia maligna di questo organo . Sottotipi di tessuti GBCS tra cui adenocarcinoma (AC), carcinoma a cellule squamose (SCC) e carcinoma adenosquamoso (ASC) sono stati esaminati su TMA mediante colorazione immunoistochimica con un anticorpo CD109. La colorazione CD109 è risultata negativa in tutti i tessuti normali della cistifellea e nei tessuti AC. Nel frattempo, le cellule positive CD109 sono state trovate nell ‘ 86,7% dei SCC e nel 91,7% degli ASC. Come CD109 è distintamente espresso in cellule sqamous maligne nella cistifellea, CD109 può essere un marcatore diagnostico per SCCS cistifellea e ASCs .
Via di segnalazione TGF-β e SCCs
La via di segnalazione TGF-β è coinvolta in molti processi cellulari tra cui la crescita cellulare, l’apoptosi di differenziazione cellulare e l’omeostasi cellulare . La famiglia di ligandi TGF-β, TGF-β1, TGF-β2 e TGF-β3, si lega ai recettori specifici della serina/treonina chinasi transmembrana di tipo I e di tipo II (TGF-ßR1 e TGF-ßR2), con conseguente attivazione dell’attività della chinasi TGF-ßR1 . Il TGF-ßR1 attivato poi propaga il segnale fosforilando i suoi substrati intracellulari, R-SMADs (Smad2 e Smad3) . Smad2 e Smad3 interagiscono con TGF-ßR1 e SARA (ancoraggio Smad per l’attivazione del recettore), una proteina del dominio FYVE che interagisce direttamente con Smad2 e Smad3, funzioni di SARA per reclutare Smad2 al recettore TGF-β , quindi i complessi eteromerici R-SMAD fosforilati formano con Co-SMAD (Smad4) . Dopo la fosforilazione e i successivi complessi con Smad4, questi complessi R-Smads vengono rilasciati da TGF-ßR1 e SARA , quindi traslocano nel nucleo dove interagiscono con fattori di trascrizione che li reclutano in specifici elementi promotori di geni bersaglio .
L’endocitosi recettoriale è un meccanismo di regolazione fondamentale nella trasduzione del segnale. I recettori TGF-β sono interiorizzati tramite vie dipendenti da clatrina e caveole. L’internalizzazione dei recettori TGF-β attraverso le fosse rivestite di clatrina è stata collegata alla segnalazione tramite Smad2/3 e al riciclaggio dei recettori. Al contrario, la localizzazione del recettore TGF-β nelle caveole è associata alla downregolazione della segnalazione Smad2/3 e alla degradazione del recettore dopo l’ubiquitinazione da parte della ligasi E3-ubiquitina Smurf2 . Tuttavia, gli Smad inibitori (Smad6 e Smad7) formano una sottoclasse distinta di Smad che agiscono in modo opposto agli R-Smad e antagonizzano la segnalazione . Possono competere con R-Smads per il legame con TGF-ßR1 attivato e quindi per inibire la fosforilazione di R-Smads . Inoltre, reclutano ligasi E3-ubiquitina al TGF-ßR1 attivato, con conseguente ubiquitinazione e degradazione del recettore e terminazione della segnalazione .
La disregolazione della via TGF-β è stata implicata in diversi tipi di cancro . Gli studi hanno dimostrato che la segnalazione TGF-β suscita un effetto preventivo durante le fasi precedenti della tumorigenesi, ma un effetto soppressivo durante il successivo sviluppo del tumore . Mutazioni nel gene TGF-ßR1 sono state trovate anche in SCC della pelle, suggerendo che l’inattivazione di TGF-β porta all’inizio di SCC .
CD109 e TGF-β via di segnalazione
CD109 è un co-recettore TGF-β, e modula TGF-β attività del recettore di segnalazione in un modo cellulare specifico . Sulla superficie cellulare, CD109 regola negativamente la via di segnalazione TGF-β1 attraverso la formazione di un complesso recettoriale con TGF-ßR1 e TGF-ßR2 nei cheratinociti umani . I recettori TGF-β degradano dopo l’ubiquitinazione da parte della E3-ubiquitina ligasi Smurf2 e sono interiorizzati tramite vie dipendenti dalla clatrina e dalle caveole. Bizet et al. dimostrato che CD109 si associa a caveolin-1 e promuove l’endocitosi del recettore TGF-β. Inoltre, CD109 promuove la localizzazione dei recettori TGF-β nel compartimento caveolare in presenza di ligando e facilita la degradazione del recettore TGF-β. CD109 regola anche la localizzazione e l’associazione di Smad7 / Smurf2 con TGF-ßR1. Gli effetti inibitori di CD109 richiedono l’espressione Smad7 e l’attività della ligasi ubiquitina Smurf2 . Inoltre, CD109 può essere rilasciato dalla superficie cellulare da lipasi cellulari come fosfatidilinositolo-specifica fosfolipasi C (PI-PLC). La forma solubile di CD109 conserva la sua capacità di legare TGF-β1 e confiscarlo lontano dai recettori TGF-β .
Tuttavia, Vorstenbosch et al. ha riferito che CD109 differenzialmente regolato TGF-β-indotta ALK1-Smad1/5 contro ALK5-Smad2/3 vie(ALK1 e ALK5 sono tutti TGF-β tipo I recettori). Hanno scoperto che la segnalazione TGF-β inibisce la proliferazione e la migrazione delle cellule endoteliali, mentre la segnalazione TGF-β induce anche questi processi tramite ALK1-Smad1/5 . Dimostrano che l’ALK1 è espresso e co-localizza con CD109 nei cheratinociti del topo e che i topi che sovraesprimono CD109 nell’epidermide mostrano una maggiore segnalazione ALK1-Smad1/5, ma una diminuzione della segnalazione ALK5-Smad2/3 .
Inoltre, TGF-β1 è un potente inibitore della crescita nella maggior parte delle cellule epiteliali . Hagiwara et al. ha dimostrato che la sovraespressione orale delle linee cellulari SCC CD109 ha accelerato la proliferazione cellulare e ha compromesso l’effetto anti-proliferativo mediato da TGF-β1. Al contrario, le cellule SCC con abbattimento CD109 hanno mostrato una crescita cellulare più lenta . Un alto livello di espressione CD109 ha inibito la fosforilazione Smad2, quindi attenuato la segnalazione TGF-β1 / Smad2 e altera la soppressione mediata da TGF-β1 della crescita cellulare, CD109 knockdown ha aumentato la fosforilazione Smad2 mediante stimolazione TGF-β1 . Sebbene CD109 regoli anche la segnalazione Smad1 / 5, non è stato collegato allo sviluppo di SCC. Insieme, CD109 facilita lo sviluppo di SCC attraverso l’inibizione della via TGF-β-Smad2 / 3 (Fig. 2).

Segnalazione CD109 e TGF-β. TGF-β lega il TGF-ßR2, che recluta e fosforila il TGF-ßR1. TGF-ßR1 attivato propaga il segnale a valle fosforilando direttamente Smad2 e Smad3. Gli R-SMAD fosforilati formano quindi complessi eteromerici con Smad4, combinati con fattori di trascrizione, regolano la trascrizione genica e la funzione cellulare. I recettori TGF-β interiorizzano attraverso le fosse rivestite di clatrina o la via caveolare. CD109 aumenta il legame TGF-β ai recettori TGF-β e promuove la localizzazione del recettore TGF-β alle caveole, in definitiva aumenta l’endocitosi del recettore TGF-β e facilita la degradazione del recettore TGF-β. Gli effetti inibitori di CD109 richiedono Smad7 espressione e Smurf2 attività della ligasi di ubiquitin
CD109 e STAT3 segnalazione
trasduttore di Segnale e attivatore del fattore di trascrizione 3 (STAT3) è fondamentale per la proliferazione cellulare, la differenziazione, la migrazione, la sopravvivenza, e l’oncogenesi . Litvinov et al. ha riferito che l’espressione della proteina CD109 era marcatamente diminuita nell’epidermide psoriasica rispetto alla pelle non coinvolta adiacente. Tuttavia, l’espressione di mRNA CD109 è invariata nelle placche psoriasiche rispetto alla pelle normale, suggerendo una possibilità che il rilascio di proteine CD109 sia aumentato nei cheratinociti psoriasici . Hanno suggerito che CD109 rilasciato / solubile è in grado di indurre cambiamenti molecolari che sono noti per verificarsi nella psoriasi . In vitro, hanno scoperto che la trasfezione di CD109 siRNA down-regola STAT3, rilascio di CD109 dalla superficie cellulare di cheratinociti umani in coltura. Inoltre, CD109 esogeno / ricombinante induce la segnalazione STAT3 nei cheratinociti umani . Inoltre, Chuang et al. ha riferito che l’espressione di CD109 è stata drammaticamente sovraregolata nelle cellule di adenocarcinoma polmonare metastatico e le cellule che esprimono uno shRNA CD109 (shCD109) hanno mostrato una drastica riduzione della fosforilazione STAT3. Il knockdown STAT3 ha notevolmente ridotto le metastasi e il ripristino dell’attività STAT3 ha aumentato la capacità delle cellule che esprimono shCD109 di metastatizzare . Al momento dell’attivazione, STAT3 è fosforilato dalla proteina non recettoriale tirosin chinasi janus chinasi 2 (JAK2), portando alla formazione di STAT3 dimero e traslocazione nel nucleo . Tuttavia, l’inibizione dell’attività della chinasi di JAK nei fibroblasti che sovraesprimono CD109 ha ridotto lo STAT3 fosforilato a un livello simile a quello delle cellule parentali che esprimono bassi livelli di CD109, suggerendo che la fosforilazione dello STAT3 indotta da CD109 richiede l’attività della chinasi di JAK. Pertanto, la segnalazione JAK / STAT3 potrebbe mediare gli effetti del CD109 nella crescita del tumore e nelle metastasi .
Sebbene l’abbattimento del CD109 nei cheratinociti umani e nelle cellule di adenocarcinoma polmonare downregulates STAT3 signaling in vitro , i topi CD109-carenti hanno mostrato risultati opposti. Mii et al. topi CD109-carenti generati, che hanno esibito le anomalie della pelle compreso l’iperplasia epiteliale e l’infiltrazione infiammatoria delle cellule. Hanno riferito che la fosforilazione STAT3 nei topi carenti di CD109 era significativamente più alta rispetto ai topi di tipo selvaggio. Inoltre, l’up-regulation della segnalazione STAT3 è associata ad una maggiore proliferazione e ad una ridotta differenziazione dei cheratinociti .
La discrepanza dei risultati degli studi in vitro e in vivo potrebbe essere causata dai cambiamenti sistematici del microambiente nei tessuti di topi con deficit di CD109. La perdita di CD109 in tutte le cellule nei topi potrebbe modificare il microambiente sottocutaneo che attiva la segnalazione STAT3 nei cheratinociti. Inoltre, CD109 può esercitare effetti regolatori distinti in diversi tipi di cellule, portando a modifiche specifiche di tipo cellulare nella segnalazione STAT3. Oltre ai cheratinociti, CD109 è espresso in cellule endoteliali, cellule epiteliali e fibroblasti, che partecipano costituiscono il tessuto cutaneo. Tuttavia, ad oggi la relazione di segnalazione CD109 e STAT3 non è stata esplorata in questi tipi di cellule.
CD109 e EGFR segnalazione
Epidermal growth factor receptor (EGFR) è un membro della famiglia di recettori ErbB. Su legante legante da EGF, EGFR forma dimeri, omodimeri o eterodimeri con un altro membro della famiglia HER2 ErbB . I recettori dimerizzati si auto-fosforilano l’un l’altro e quindi fosforilano la proteina non recettoriale tirosina c-Src chinasi, che attiva STAT3 . L’attivazione di EGFR promuove la migrazione cellulare, la sopravvivenza e la proliferazione. Nei tumori maligni EGFR over-espressione è correlata con la profondità di invasione del tumore e legata alla prognosi più povera . Le mutazioni che portano alla sovraespressione di EGFR sono rilevate in SCC polmonare , SCC testa e collo e SCC esofago . Il CD109 ancorato alla membrana nelle cellule SK-MG-1 interagisce direttamente con EGFR e migliora la segnalazione EGF, che successivamente aumenta la migrazione e l’invasione cellulare, mentre il CD109 secreto non ha alcun effetto sulla segnalazione EGF . EGFR potrebbe mediare gli effetti di CD109 sulla segnalazione STAT3, che richiede ulteriori studi per chiarire (Fig. 3).
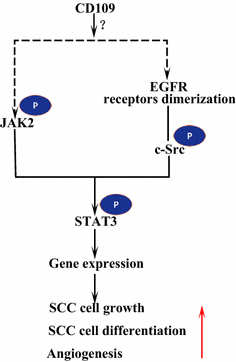
CD109 e EGFR / STAT3 segnalazione. CD109 facilita la segnalazione EGFR. I recettori dimerizzati di EGFR fosforilano c-Src, che a sua volta fosforila STAT3. CD109 può anche attiva STAT3 attraverso la fosforilazione di JAK2, quindi promuovere la crescita delle cellule SCC, differenziazione e angiogenesi
Ad oggi il quadro generale della tumorigenesi e lo sviluppo di SCC rimangono incompleti. Esistono colloqui incrociati tra TGF-β e vie di segnalazione STAT3 o EGFR. CD109 è probabilmente uno degli effettori chiave della rete di segnalazione che regola SCC. Tuttavia, nessuna prova diretta è stata riferita per delineare i ruoli di CD109 nella patogenesi di SCCs.
Prospettive
Gli studi effettuati su campioni di tessuto umano indicano che il CD109 è altamente espresso in SCC di organi multipli, in particolare in cellule squamose maligne ben differenziate . Rilevando l’espressione di CD109 con immunoistochimica nei tessuti umani, CD109 può potenzialmente agire come biomarcatore per determinare la progressione di SCC. Gli studi attuali suggeriscono che CD109 è altamente espresso in SCC ben differenziati e la sua espressione è inferiore in SCC non differenziati . Tuttavia, non è chiaro se CD109 sia associato a invasione vascolare, metastasi e prognosi dopo l’intervento chirurgico. Pertanto, sono necessari ulteriori studi per esplorare il significato clinico-patologico di CD109 in SCC in un campione di dimensioni maggiori.
CD109 è una glicoproteina legata al GPI, che gli consente di essere rilasciata dalla membrana . La forma solubile di CD109 inoltre pregiudica il legame di TGF-β ai suoi ricevitori e successivamente modula la progressione di SCC . Litvinov et al. trovato che CD109 rilasciato dalla superficie cellulare nell’ambiente extracellulare, e la forma rilasciata di CD109 mantiene la sua capacità di indurre vie di segnalazione intracellulari. Inoltre, Sakakura et al. ha riferito che il siero CD109 è stato rilasciato dal tumore xenografted e aumenta proporzionalmente con il volume di xenotrapianto tumorale. Pertanto, il rilevamento del livello sierico di CD109 potrebbe aiutare a monitorare i tumori che sovraesprimono CD109 incluso SCC.
Gli esosomi comunicano la lesione tumorale primaria e la sua nicchia attraverso il suo pacchetto contenente proteine selezionate o altre molecole . L’isolamento e le analisi dell’esosoma associato al tumore circolante possono servire da biomarcatori per la diagnosi dei pazienti oncologici. CD109 esogeno è stato identificato come componente dell’esosoma secreto da 293 cellule trasfettate, rendendolo un bersaglio promettente per la diagnosi dell’esosoma . Ancora, fino ad oggi CD109 non sono stati riportati in esosomi derivati da SCC o altre cellule tumorali. I meccanismi alla base del confezionamento di CD109 negli esosomi meritano un’indagine più dettagliata.
Oltre al potenziale come biomarcatore, CD109 potrebbe essere un bersaglio per approcci terapeutici. CD109 è una proteina di membrana, che può essere mirata direttamente da anticorpi o enzimi specifici. CD109 potrebbe anche essere riconosciuto da veicoli mirati per la consegna di droga. Tuttavia, i ruoli dettagliati di CD109 nella patogenesi di SCC non sono ancora chiari. Ad esempio, è necessario definire la relazione di espressione inferiore di CD109 e SCC indifferenziati. L’intervento di CD109 può essere preso in considerazione nella pratica clinica solo dopo che i rischi e i benefici sono stati valutati attentamente.