
a laphámsejtes carcinoma (SCC), más néven prickle cell carcinoma, a rákkal kapcsolatos halálozás egyik vezető oka világszerte. Az SCC egy rosszindulatú epitheliális daganat, amely olyan szövetekben keletkezik, amelyek gátat képeznek a szervezet és a környezet között, mint például a bőr, a szájüreg, az üreg, a nyelőcső és a tüdő . Egy korábbi szakaszban az SCC-t epidermális keratinizáció és fekélyképződés kíséri a nyálkahártya felületén, és a mélyebb szöveteket egy későbbi szakaszban az SCC-k támadják meg. A mikroszkopikus megjelenés érdekében az SCC sejtek fészkekbe rendeződnek, amelyeket sokszögű sejtek vesznek körül, külön sejthatárokkal és hiperkromatikus magokkal. Az SCC-k hajlamosak a metasztázisra, általában a regionális nyirokcsomókon keresztül, és több szerv szisztematikus károsodását okozhatják. A sebészet, a lézerterápia és a sugárzás továbbra is a legszélesebb körben alkalmazott kezelés az SCCs számára . Az SCC variánsok egyedi citomorfológiai jellemzői különböző kezelésekhez és eredményekhez vezetnek .
a CD109 sejtfelszíni antigén egy glikozil-foszfatidilinozit (GPI)-kapcsolt glikoprotein, körülbelül 170 kDa, és a tioésztert tartalmazó fehérjék A2 makroglobulin (a2M) / C3, C4, C5 családjának tagja . Az emberi CD109 gén a 6Q kromoszómában található, amely a teljes CD109 genomi szekvencia körülbelül 3,3% – át teszi ki 33 exonjával . Az izolált CD109 cDNS egy 4335 bp nyílt leolvasású keretet tartalmaz, amely 1445 aminosavat (aa) kódol . A CD109 fehérje tartalmaz egy 21 aa N-terminális leader peptidet, egy feltételezett csali régiót (aa 651-683), egy tioészter kötőhelyet (aa 918-924), egy tioészter reaktivitást meghatározó hexapeptidet (aa1030–1035) és egy C-terminális konszenzus GPI-anchor jelsorozatot, amelynek hasítása az 1420 aminosav után várható (ábra. 1) . Az N-terminális vezető peptid a fehérjét a periplazmán belüli belső membránhoz rögzíti, valamint a cd109 N-terminális fragmentumát, amelyet a Furin proteáz hasítása után választanak ki a sejtekből . A nagyrészt rendezetlen csali régió proteáz hasítása aktiválja a CD109-et, ami konformációs változást eredményez, amely csapdába ejti a proteázt egy ketrecszerű szerkezetben, és kiteszi a nagyon reaktív tioészter kötést. A tioészter kötőhelye, amely aktiváláskor kovalensen összekapcsolhatja a CD109-et a támadó proteáz felületén lévő lizinmaradványokkal. Ezenkívül a C-terminális konszenzus GPI-horgony jelszekvenciáját receptorkötő doménnek is nevezik .

The structure of CD109 protein. A CD109 fehérje teljes hossza 1445 aminosavból (aa) áll, beleértve egy 21 aa N-terminális szignálpeptidet, egy feltételezett csali régiót (aa 651-683), egy tioészter kötőhelyet (aa 918-924), egy tioészter reaktivitást meghatározó hexapeptidet (aa1030-1035) és egy C–terminális konszenzus GPI-anchor jelsorozatot (aa 1420-1445), amelynek hasítása az aminosav után várható 1420
a cd109-et először a primitív limfoid/mieloid sejtvonal ellen emelt monoklonális antitest azonosította sejtfelszíni antigénként kg1a . Korábbi tanulmányok arról számoltak be, hogy a CD109 a magzati és felnőtt CD34+ pozitív csontvelő mononukleáris sejtek, az aktivált T-limfoblasztok, az aktivált vérlemezkék, az endothel sejtek, a mesenchymalis őssejt-alcsoportok és számos emberi tumorsejt-vonal egy részhalmazában expresszálódik , de nem expresszálódik nyugvó emberi T-sejtekben, vérlemezkékben vagy perifériás vér leukocitákban . Különösen a CD109-et mutatták ki több szerv SCCs-jében, beleértve a tüdőt , a nyelőcsövet , a méhnyakot , a bőrt , a péniszt, a szájüreget és az epehólyagot . A CD109 a transzformáló növekedési faktor (transzformáló növekedési faktor) (TGF-61) receptorrendszer egyik összetevője, és negatívan szabályozza a TGF-1 jelátvitelt . A funkcionális vizsgálatok vesztesége és nyeresége azt sugallta, hogy a CD109 molekuláris célpont lehet a különböző szöveti eredetű SCC-k új terápiáinak kifejlesztéséhez . Ebben az áttekintésben összefoglaljuk a CD109 jelenlegi ismereteit és az SCCs-hez fűződő kapcsolatát.
CD109 expresszió az SCCs-ben
a tumorszövetekben a CD109 immunhisztokémiailag kimutatható volt az SCCs-ben, valamint urothelialis carcinomákban , malignus melanomákban , bazális-szerű emlő carcinomákban , myxofibrosarcomában , epithelialis sarcomákban és gliomában . Különösen, Shiraki et al. jelentett CD109-pozitív perivaszkuláris tumorsejtek humán alacsonyabb fokú glioma szövetekben és egy egér modellben összefoglalt humán glioma, ami arra utal, hogy a CD109 kulcsszerepet játszik ebben a betegségben. Korábbi tanulmányok kimutatták, hogy a CD109 magas expressziója az SCCs-ben és a korlátozott expresszió a normál laphámsejtekben (1 .táblázat). Ezenkívül a CD109 erősen expresszálódik jól differenciált SCC-kben, nem pedig közepesen vagy rosszul differenciált SCC-kben, így a CD109 expressziós szintje fordítottan korrelál a tumor fokozatával .
CD109 és lung SCC
a tüdő laphámsejtes karcinóma (LSCC) a nem kissejtes karcinómák egyik fő altípusa-sejtes tüdőrák (NSCLC), és az NSCLC-k eseteinek körülbelül 20-30% – át teszi ki. A CD109 expressziót humán tüdősejtes karcinómákban kvantitatív RT-PCR-rel vizsgáltuk, amely szignifikánsan nagyobb cd109 expressziót mutatott laphámsejtes karcinómákban, de nem adenokarcinómákban, nagysejtes karcinómákban vagy kissejtes karcinómákban . Továbbá, Sato et al. CD109 antitestet használt a CD109 expressziójának elemzésére normál tüdő-és tüdőkarcinóma szövetekben. A normál tüdőszövetekben a CD109 expresszió elsősorban a bronchiális és bronchiolaris hámsejtek bazális sejtjeire korlátozódott . A tüdőkarcinóma szöveteiben elsősorban az Lscc-kben mutattak ki erős immunreaktivitást, de más típusú tüdőkarcinómákban nem . Ezek az eredmények azt mutatják, hogy a CD109 kifejezetten fel van szabályozva az LSCCs – ben.
CD109 és a nyelőcső SCC
a nyelőcső laphámsejtes karcinóma (ESCC) az összes rákkal kapcsolatos halálozás egyhatodát teszi ki, magasabb incidenciával fordul elő az ázsiai országokban . Az ESCC prognózisa továbbra is gyenge, az ESCC 5 éves túlélési aránya körülbelül 14-22% . A legújabb tanulmányok azt sugallják, hogy a TGF-6CC receptorok megváltozott expressziója hozzájárul az ESCC progressziójához, a TGF-CC gátló fehérjéinek emelkedett expressziója pedig a 6CC jelátvitel korrelál az ESCC rossz prognózisával . Hashimoto et al. a CD109 expressziót az ESCC szövetekben kvantitatív RT-PCR-rel vizsgálták, és a CD109 mRNS transzkripciót az Escc-k körülbelül 50% – ában szabályozták. Később a CD109 fehérje szintjét az ESCC-ben immunhisztokémiával vizsgáltuk szöveti mikroáramkörökön (TMA) . A nem beteg nyelőcsőszövetben a CD109 expressziója korlátozott volt a rétegzett hámsejtek citoszoljában gyenge szinten. A TMA-k összes ESCC-mintájában a rosszindulatú laphámsejtek szálai erős CD109 festést mutattak a citoszolban. Más sejttípusokban nem figyeltek meg CD109 pozitív festést. Ezenkívül a CD109 expressziója magasabb volt a jól és közepesen differenciált Escc – kben, mint a rosszul differenciáltakban .
CD109 és cervix SCC
a cervix (cervix) rák a nők rákkal összefüggő halálának körülbelül 12%-át teszi ki . Az SCCs az összes méhnyakrák 80-85% – át teszi ki . Zhang et al. kimutatta, hogy a CD109 expresszió szignifikánsan magasabb volt a nyaki SCC-kben, mint az endometrium adenokarcinómáiban. Megvizsgálták a CD109 expressziót öt humán méhnyakrák sejtvonalban, és megfigyelték a CD109 expresszió magas szintjét két SCC sejtvonalban.
CD109 és cutan SCC
a cutan squamous cell carcinoma (CSCC) a nem melanoma bőrrák (nmsc) második leggyakoribb típusa, folyamatosan növekvő előfordulási gyakorisággal . Bár a CSCC-nek általában kedvező prognózisa van, még mindig a betegek körülbelül 1,5–2% – a hal meg ebben a betegségben . A CD109 expressziót a CSCCs-ben immunhisztokémiával vizsgálták a TMA-kon . A normál bőrben a CD109 gyengén expresszálódott az epidermális sejtek bazális rétegében, míg a rosszindulatú pikkelyes epidermális sejtek szálai erős CD109 festést mutattak. Ezenkívül a CD109 expresszió fordítottan korrelált a CSCC osztályzatokkal. Az Escc – khez hasonlóan a CD109 expressziója is magasabb volt a jól és közepesen differenciált Cscc-kben, mint a rosszul differenciáltakban . A Penile squamous cell carcinoma (PSCC) a Cscc-k egyik altípusa, és ritkán tanulmányozták . Dong et al. kimutatták, hogy a CD109 fehérje erősen expresszálódik a Pscc-k rosszindulatú laphámsejtjeiben, összehasonlítva a normál péniszszövetekkel a TMA-kon. Továbbá az expressziós mintát friss műtéti PSCC mintákon validálják immunfluoreszcenciával, qRT-PCR-rel és western blott-tal, ami arra utal, hogy a CD109 lehet A pscc biomarkere .
CD109 és szájüregi SCC
az orális laphámsejtes karcinóma (OSCC) az összes rák 2-3% – át teszi ki világszerte . A funkciómutáció elvesztése a TGF-KB II típusú receptorban gyakori esemény a szájüreg SCC-jében . A CD109 expresszióját 124 beteg normál orális szöveteiben és OSCC szöveteiben immunhisztokémiai festéssel vizsgálták. A CD109 expresszió magas szintjét gyakran észlelték az SCCs-ben és a szájüreg premalignus elváltozásaiban, de a normál laphám hámban nem . Ezenkívül a CD109 expressziós szintje szignifikánsan magasabb volt a jól differenciált Oscc-kben, mint a közepesen vagy rosszul differenciált Oscc-kben, ami azt jelenti, hogy a CD109 expresszió korrelál az Oscc-k differenciálási szakaszaival . Ezenkívül a CD109-et túlexpresszáló OSCC sejtvonalak gyorsított sejtnövekedést mutattak in vitro, ami arra utal, hogy a CD109 részt vesz az OSCCs progressziójában.
CD109 és epehólyag SCC
az epehólyagrák (GBC) a leginkább agresszív az eperákok közül, a legrövidebb medián túléléssel . Az epehólyag laphámsejtes karcinóma (GSCC) a szerv rosszindulatú daganatának 3% – át teszi ki . A GBC-szövetek altípusait, köztük az adenocarcinomát (AC), a laphámsejtes karcinómát (SCC) és az adenosquamous carcinomát (ASC) TMA-kkal vizsgálták CD109 antitesttel végzett immunhisztokémiai festéssel. A CD109 festés negatív volt minden normális epehólyag szövetben és AC szövetben. Eközben CD109 pozitív sejteket találtak az SCC-k 86,7% – ában és az ASC-k 91,7% – ában. Mivel a CD109 egyértelműen expresszálódik az epehólyag rosszindulatú sqamous sejtjeiben, a CD109 diagnosztikai marker lehet az epehólyag SCCs és ASC-k számára .
TGF-ons jelátviteli út és SCCs
a TGF-CA jelátviteli út számos sejtfolyamatban részt vesz, beleértve a sejtnövekedést, a sejtdifferenciálódási apoptózist és a sejt homeosztázisát . A család, a TGF-β ligandumok, TGF-β1, TGF-β2, valamint TGF-β3, kötődik konkrét transzmembrán i., illetve II-es típusú szerin/treonin kináz receptor (TGF-ßR1, valamint TGF-ßR2) , ami aktiválás TGF-ßR1 kináz aktivitás . Ezután az aktivált TGF-xhamster1 az intracelluláris szubsztrátok, az R-SMAD-ok (Smad2 és Smad3) foszforilálásával terjeszti a jelet . Az Smad2 és az Smad3 kölcsönhatásba lép a TGF-xhamster1-gyel és a SARA-val (SMAD anchor for receptor activation), egy FYVE domén fehérjével, amely közvetlenül kölcsönhatásba lép az Smad2-vel és az Smad3-mal , A SARA feladata, hogy az Smad2-t a TGF-ons receptorhoz Gyűjtse, majd a foszforilált R-SMAD-ok heteromer komplexeket képeznek a Co-SMAD-val (Smad4) . Az Smad4-gyel végzett foszforiláció és az azt követő komplexek után ezek az R-SMAD-komplexek felszabadulnak a TGF-xhamster1-ből és a SARA-ból, majd transzlokálódnak a magba , ahol kölcsönhatásba lépnek a transzkripciós faktorokkal, amelyek a célgének specifikus promoter elemeihez toborozzák őket .
A Receptor endocitózis kulcsfontosságú szabályozó mechanizmus a jelátvitelben. A TGF-ons receptorok mind a clathrin -, mind a caveolae-függő utakon keresztül internalizálódnak. A clathrin-bevonatú gödrökön keresztül a TGF-ons receptorok internalizálása összekapcsolódott az Smad2/3-on keresztüli jelátvitellel és a receptor újrahasznosításával. Ezzel szemben a caveolae-ban a TGF-6 receptor lokalizációja a Smad2/3 jelátvitel és a receptor degradációjának csökkenésével jár együtt az E3-ubiquitin ligáz Smurf2 általi ubiquitinációt követően . Azonban a gátló SMAD-ok (Smad6 és Smad7) a SMAD-ok egy külön alosztályát alkotják, amelyek ellentétes módon hatnak az R-SMAD-okkal és antagonizálják a jelátvitelt . Versenyezhetnek az R-SMAD-okkal az aktivált TGF-kb-1-hez való kötődésért, és így gátolhatják az R-SMAD-ok foszforilációját . Ezen felül E3-ubiquitin ligázokat toboroznak az aktivált TGF-aponkr1-be, ami a receptor ubiquitinációját és lebomlását, valamint a jelátvitel megszűnését eredményezi .
a TGF-6 út Diszregulációja a rák több típusában is szerepet játszott . Tanulmányok kimutatták, hogy a TGF-6 jelátvitel megelőző hatást vált ki a tumorigenezis korábbi szakaszaiban, de szuppresszív hatást vált ki a tumor későbbi fejlődése során . A bőr SCCS-jében is találtak mutációkat a TGF-intravitr1 génben, ami arra utal, hogy a TGF inaktiválása-a CA-k inaktiválása az SCC-k inaktiválásához vezet .
CD109 és TGF-jelátviteli út
a CD109 egy TGF-CAC-Ko-receptor , és a TGF-jelátviteli receptor aktivitását sejtspecifikus módon modulálja . A sejtfelszínen a CD109 negatívan szabályozza a TGF-ca-1 jelátviteli útvonalat azáltal, hogy humán keratinocitákban TGF-a-R1 és TGF-a-R2 receptor komplexet képez . A TGF-6 receptorok az E3-ubiquitin ligáz Smurf2 általi ubiquitinációt követően bomlanak le, és mind a clathrin-függő, mind a caveolae-függő utakon internalizálódnak . Bizet et al. kimutatták, hogy a CD109 társul a caveolin-1-hez, és elősegíti a TGF-6 receptor endocitózist. Ezenkívül a CD109 ligandum jelenlétében elősegíti a TGF-adapterek lokalizációját a caveoláris kompartmentbe, és megkönnyíti a TGF-adapterek lebomlását. A CD109 szabályozza a Smad7/Smurf2 lokalizációját és társítását a TGF-xhamster1-gyel. A CD109 gátló hatásai Smad7 expressziót és Smurf2 ubiquitin ligáz aktivitást igényelnek . Ezenkívül a CD109 felszabadulhat a sejtfelszínről celluláris lipázok, például foszfatidilinozit-specifikus foszfolipáz C (PI-PLC). A CD109 oldható formája megtartja a képességét, hogy megköti a TGF-xhamster1-et, és elkobozza a TGF-6 receptoroktól .
azonban Vorstenbosch et al. beszámoltak arról, hogy a CD109 differenciáltan szabályozza a TGF-indukált alk1-Smad1 / 5-et az ALK5-Smad2/3 útvonalakkal szemben (az ALK1 és az ALK5 mind TGF-6 I. típusú receptorok). Azt találták, hogy a TGF-β jelzés gátolja az endothel sejt proliferációt, valamint átállási, míg a TGF-β jelzés is indukálja ezek a folyamatok keresztül ALK1-Smad1/5 . Kimutatták, hogy az ALK1 expresszálódik és együtt lokalizálódik a CD109-gyel az egér keratinocitáiban, és hogy az epidermiszben a CD109-et túlexpresszáló egerek fokozott ALK1-Smad1/5 jelátvitelt mutatnak, de csökkent ALK5-Smad2/3 jelátvitelt mutatnak .
különben is, a TGF-xhamster1 a növekedés erős inhibitora a legtöbb hámsejtben . Hagiwara et al. kimutatták, hogy az orális SCC sejtvonalak túlzott expressziója a CD109 felgyorsította a sejtproliferációt és rontotta a TGF-6 által közvetített anti-proliferatív hatást. Ezzel szemben a CD109 leütéssel rendelkező SCC sejtek lassabb sejtnövekedést mutattak . A CD109 expresszió magas szintje gátolta a Smad2 foszforilációt, így gyengítette a TGF-61/Smad2 jelátvitelt, és rontotta a sejtnövekedés TGF-1 által közvetített szuppresszióját, a CD109 leütése fokozta az Smad2 foszforilációját a TGF-61 stimulációval . Bár a CD109 szabályozza az Smad1 / 5 jelzést is , nem kapcsolódik az SCC fejlesztéséhez. A CD109 együttesen megkönnyíti az SCC-k kialakulását a TGF-6-smad2/3 út gátlása révén (ábra. 2).

CD109 és TGF-jelek. A TGF-xhamsterhez kötődik a TGF-xhamster2, amely a TGF-xhamster1-et veszi fel és foszforilálja. Az aktivált TGF-enterpr1 az smad2 és Smad3 közvetlen foszforilezésével továbbterjeszti a jelet. A foszforilált R-SMAD-ok ezután heteromer komplexeket képeznek az Smad4-gyel, transzkripciós faktorokkal kombinálva szabályozzák a gén transzkripcióját és a sejtek működését. A TGF-6 receptorok a clathrin-bevonatú gödrökön vagy a caveolar útvonalon keresztül internalizálódnak. CD109 növeli a TGF-β kötelező TGF-β-receptorok, valamint elősegíti a TGF-β-receptor lokalizáció, hogy a caveolae, végső soron növeli a TGF-β-receptor endocytosis, valamint elősegíti a TGF-β-receptor lebomlás. A CD109 gátló hatásai Smad7 expressziót és Smurf2 ubiquitin ligáz aktivitást igényelnek
a CD109 és a STAT3 jelátvitel
a transzkripciós faktor 3 (STAT3) Jelátalakítója és aktivátora kritikus a sejtproliferáció, a differenciálódás, a migráció, a túlélés és az onkogenezis szempontjából . Litvinov et al. arról számoltak be, hogy a CD109 fehérje expressziója jelentősen csökkent a pszoriázisos epidermiszben a szomszédos, nem érintett bőrhöz képest. A CD109 mRNS expressziója azonban változatlan a pszoriázisos plakkokban a normál bőrhöz képest, ami arra utal, hogy a CD109 fehérje felszabadulása fokozódik a pszoriázisos keratinocitákban . Azt javasolták, hogy a felszabadult / oldható CD109 képes molekuláris változásokat indukálni, amelyekről ismert, hogy psoriasisban fordulnak elő . In vitro azt találták, hogy a CD109 siRNS transzfekciója lefelé szabályozza a STAT3-ot, a CD109 felszabadulását a tenyésztett humán keratinociták sejtfelszínéről. Ezenkívül az exogén / rekombináns CD109 STAT3 jelátvitelt indukál az emberi keratinocitákban . Különben is, Chuang et al. arról számoltak be, hogy a CD109 expresszió drámai módon szabályozódott a metasztatikus tüdő adenokarcinóma sejtekben, és a CD109 shRNS-t (shCD109) expresszáló sejtek drámai csökkenést mutattak STAT3 foszforiláció. A STAT3 leütése jelentősen csökkentette az áttéteket, a STAT3 aktivitás helyreállítása pedig növelte az shCD109-expresszáló sejtek metasztázisképességét . Aktiválás után a STAT3-at a nem receptor fehérje, a tirozin-kináz janus-kináz 2 (JAK2) foszforilálja, ami STAT3 dimer képződéséhez és a sejtmagba történő transzlokációhoz vezet . A CD109-et túlexpresszáló fibroblasztokban a jak-kináz aktivitás gátlása azonban a foszforilezett STAT3-t hasonló szintre csökkentette, mint az alacsony CD109-szintet expresszáló szülői sejtekben, ami arra utal, hogy a CD109 által indukált STAT3-foszforiláció jak-kináz aktivitást igényel. Így a JAK / STAT3 jelátvitel közvetítheti a CD109 hatását a tumor növekedésében és a metasztázisban .
bár a CD109 leütése az emberi keratinocitákban és a tüdő adenokarcinóma sejtekben in vitro csökkenti a STAT3 jelátvitelt, a CD109-hiányos egerek ellentétes eredményeket mutattak. Mii et al. generált CD109-hiányos egerek, amelyek bőrbetegségeket mutattak, beleértve az epithelialis hiperpláziát és a gyulladásos sejtek infiltrációját. Beszámoltak arról, hogy a STAT3 foszforiláció CD109-hiányos egerekben szignifikánsan magasabb volt a vad típusú egerekhez képest. Ezenkívül a STAT3 jelátvitel fokozott szabályozása a megnövekedett proliferációval és a keratinociták károsodott differenciálódásával jár .
az in vitro és in vivo vizsgálatok eredményeinek eltérését a mikrokörnyezet szisztematikus változása okozhatja a CD109 hiányos egerek szöveteiben. A CD109 elvesztése az egerek összes sejtjében módosíthatja a szubkután mikrokörnyezetet, amely aktiválja a stat3 jelátvitelt a keratinocitákban. Ezenkívül a CD109 különféle szabályozó hatásokat gyakorolhat a különböző sejttípusokban, ami a STAT3 jelátvitel sejttípus-specifikus módosulásához vezethet. Amellett, hogy a keratinociták, CD109 expresszálódik endothel sejtek, hámsejtek, fibroblasztok, amelyek részt vesznek alkotják a bőrszövet. A CD109 és a STAT3 jelátvitel kapcsolatát azonban a mai napig nem vizsgálták ezekben a sejttípusokban.
CD109 és EGFR jelátvitel
az epidermális növekedési faktor receptor (EGFR) az Erbb receptorcsalád tagja. Az EGF által kötött ligandum esetén az EGFR dimereket képez, akár homodimereket, akár heterodimereket az ErbB család másik tagjával HER2 . A dimerizált receptorok auto-foszforilálják egymást, majd foszforilálják a nem receptor fehérjét tirozin c-Src kináz, amely aktiválja STAT3 . Az EGFR aktiválása elősegíti a sejtek migrációját, túlélését és proliferációját. A rosszindulatú daganatokban az EGFR túlzott expressziója korrelál a tumor inváziójának mélységével és a rosszabb prognózishoz kapcsolódik . Az EGFR túlzott expressziójához vezető mutációkat a tüdő SCC-ben , a fej és a nyak SCC-ben és a nyelőcső SCC-ben észlelik . A membránnal rögzített CD109 az SK-MG-1 sejtekben közvetlenül kölcsönhatásba lép az EGFR-rel, és fokozza az EGF jelátvitelt, ami később növeli a sejtek migrációját és invázióját, míg a szekretált CD109 nincs hatással az EGF jelátvitelre . Az EGFR közvetítheti a CD109 STAT3 jelzésre gyakorolt hatásait, ami további vizsgálatokat igényel a tisztázáshoz (ábra. 3).
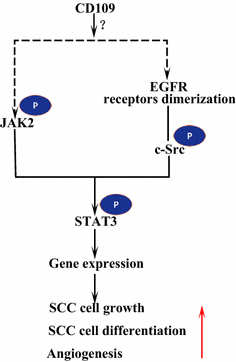
CD109 és EGFR/STAT3 jelzés. A CD109 megkönnyíti az EGFR jelzést. A dimerizált EGFR receptorok foszforilálják a c-Src-t, ami viszont foszforilálja a STAT3-ot. A CD109 a STAT3-at a JAK2 foszforilezésével is aktiválhatja, majd elősegítheti az SCC sejtek növekedését, differenciálódását és angiogenezist
a mai napig a tumorigenezisről és az SCC fejlődéséről alkotott átfogó kép hiányos. Vannak keresztbeszélgetések a TGF-KB és a STAT3 vagy EGFR jelátviteli utak között. A CD109 valószínűleg az SCCs-t szabályozó jelzőhálózat egyik legfontosabb effektora. Azonban nem jelentettek közvetlen bizonyítékot a CD109 szerepének felvázolására az SCCs patogenezisében.
perspektívák
az emberi szövetmintákból származó vizsgálatok azt mutatják, hogy a CD109 erősen expresszálódik több szerv SCCs-jében, különösen jól differenciált rosszindulatú laphámsejtekben . A CD109 expresszió kimutatásával immunhisztokémiával az emberi szövetekben a CD109 potenciálisan biomarkerként hat az SCC progressziójának meghatározására. A jelenlegi tanulmányok azt sugallják, hogy a CD109 erősen expresszálódik a jól differenciált SCCs – ben, expressziója pedig alacsonyabb a nem differenciált SCCs-ben . Nem világos azonban, hogy a CD109 összefügg-e a vaszkuláris invázióval, metasztázissal és a műtét utáni prognózissal. Ezért további vizsgálatokra van szükség a CD109 klinikai patológiai jelentőségének feltárásához az SCCs-ben nagyobb mintaméretben.
a CD109 egy GPI-hez kapcsolt glikoprotein, amely lehetővé teszi annak felszabadulását a membránból . A CD109 oldható formája szintén befolyásolja a TGF-ons receptorokhoz való kötődését, majd modulálja az SCC progresszióját . Litvinov et al. megállapította, hogy a CD109 felszabadul a sejtfelszínről az extracelluláris miliőbe, a cd109 felszabadult formája pedig megőrzi képességét intracelluláris jelátviteli utak indukálására. Különben is, Sakakura et al. arról számoltak be, hogy a szérum CD109 xenograftált tumor által szabadult fel, és arányosan növekszik a tumor xenograft térfogatával. Ezért a szérum CD109 szint kimutatása segíthet a CD109-et túl expresszáló daganatok monitorozásában, beleértve az SCCs-t is.
az exoszómák a primer tumor elváltozást és annak rését a kiválasztott fehérjéket vagy más molekulákat tartalmazó csomagon keresztül kommunikálják . A keringő tumorhoz kapcsolódó exoszóma izolálása és elemzése biomarkerként szolgálhat a rákos betegek diagnosztizálásához. Az exogén CD109-et a transzfektált 293 sejtből szekretált exoszóma komponenseként azonosították, így ígéretes célpont az exoszóma diagnózisához . Ennek ellenére a CD109-et eddig nem jelentették az SCC-ből vagy más tumorsejtekből származó exoszómákban. A CD109 exoszómákba történő csomagolásának alapjául szolgáló mechanizmusok részletesebb vizsgálatot érdemelnek.
a biomarker potenciálja mellett a CD109 lehet a terápiás megközelítések célpontja. A CD109 egy membránfehérje, amelyet közvetlenül specifikus antitestek vagy enzimek célozhatnak meg. A CD109-et a kábítószer-szállítás célzott járművei is felismerhetik. A CD109 részletes szerepe azonban az SCCs patogenezisében még mindig nem tisztázott. Például meg kell határozni a CD109 alacsonyabb expressziójának és a differenciálatlan SCCs-nek a kapcsolatát. A CD109 beavatkozás csak a klinikai gyakorlatban mérlegelhető, miután a kockázatokat és előnyöket gondosan értékelték.