
Régulation et contrôle du métabolisme chez les bactéries (page 5)
(Ce chapitre compte 5 pages)
© Kenneth Todar, PhD
La répression par catabolite
L’induction enzymatique est toujours considérée comme une forme de contrôle négatifcausel’effet de la molécule régulatrice (le répresseur actif) est de réduire ou de réduire le taux de transcription. La catabolitérépression est untype de contrôle positif de la transcription, car une protéine régulatrice affecte une augmentation (régulation à la hausse) du taux de transcription d’un opéron. Le processus a été découvert dans E. l’effet du glucose a été considéré à l’origine comme l’effet du glucose, car il a été constaté que la glycoserépressait la synthèse de certaines enzymes inductibles, même si l’inducteur de la voie était présent dans l’environnement. La découverte a été faite lors de l’étude de la régulation de l’opéron lac chez E. coli.Puisque le glucose est dégradé par des enzymes constitutives et que le lactose est initialement dégradé par des enzymes inductibles, que se passerait-il si la bactérie était cultivée en quantités limitées de glucose et de lactose? Un graphique du taux de croissance bactérienne a été réalisé dans une courbe de croissance diauxique qui a montré deux phases distinctes de croissance active (Figure 9). Au cours de la première phase de croissance exponentielle, la bactérie utilise le glucose comme source d’énergie jusqu’à ce que tout le glucose soit épuisé.Ensuite, après une phase de latence secondaire, le lactose est utilisé pendant une deuxième étape de croissance exponentielle.

Figure 9. La courbe de croissance diauxique de E. coli cultivé à des concentrations limites d’un mélange de glucose et de lactose
Pendant la période d’utilisation du glucose, le lactose n’est pas utilisé parce que les cellules sont incapables de transporter et de cliver le lactose disaccharidique.Le glucose est toujours métabolisé en premier de préférence aux autres sucres. Seulement après l’utilisation complète du glucose, le lactose est dégradé. Le lactoséopérone est réprimé même si le lactose (l’inducteur) est présent. La ration écologique est que le glucose est une meilleure source d’énergie que le lactose.son utilisation nécessite deux enzymes de moins.
Ce n’est qu’une fois le glucose épuisé que les enzymes de lactoseutilisation sont synthétisées. Le retard secondaire pendant la croissance diauxique représente le temps nécessaire à l’induction complète de l’opéron lac et à la synthèse des enzymes nécessaires à l’utilisation du lactose (lactose perméase et bêta-galactosidase).Ce n’est qu’alors que la croissance bactérienne se produit au détriment du lactose. Puisque la disponibilité du glucose réprime les enzymes pour la lactoseutilisation, ce type de répression est devenu connu sous le nom de répression par catabolite ou d’effet glucose.
Le glucose est connu pour réprimer un grand nombre d’enzymes inductibles dans de nombreuses bactéries différentes. Le glucose réprime l’induction des opérons inductibles en inhibant la synthèse de l’AMP cyclique (AMPc), une nucléotide nécessaire à l’initiation de la transcription d’un grand nombre de systèmes d’enzymes inductibles, y compris l’opéron lac.
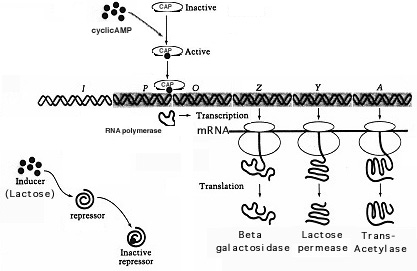
Le rôle de l’AMPc cyclique est compliqué. L’AMPc est nécessaire pouractiver une protéine allostérique appelée CAP (protéine activatrice de catabolite) qui se lie au site promoteur CAP et stimule la liaison de la RNAppolymérase au promoteur pour l’initiation de la transcription. Ainsi, pour favoriser efficacement la transcription génique de l’opéron lac, non seulement le lactose doit être présent pour inactiver le répresseur lac, mais l’AMPc doit être disponible pour se lier à CAP qui se lie à l’ADN pour faciliter la transcription. Dansla présence de glucose, l’activité de l’adénylate cyclase (AC) est bloquée. ACis requis pour synthétiser l’AMPc à partir de l’ATP. Par conséquent, si les niveaux d’AMPc sont faibles, le CAP est inactif et la transcription ne se produit pas. Dansl’absence de glucose, les niveaux d’AMPc sont élevés, le CAP est activé par l’AMPc et la transcription se produit (en présence de lactose).
De nombreux promoteurs contrôlés positivement, tels que le promoteur lac, ne sont pas pleinement fonctionnels en présence de RNAp seul et nécessitent une activation par CAP. CAP est codé par un gène régulateur distinct et présente des niveaux inconstitutifs. Le CAP n’est actif qu’en présence du Camp. L’association de l’AMPc au CAP provoque un changement conformationnel de la protéine permettant de se lier au promoteur près du site de liaison de la RNAp. CAP peut apparemment interagir avec RNAp pour augmenter le taux de transcription de l’opéron d’environ 50 fois.Le contrôle positif de l’opéron lac est illustré à la figure 10.
Figure 10. La catabolitérépression est un contrôle positif de l’opéron lac. L’effet est une augmentation du taux de transcription. Dans ce cas, la protéine CAP est activée Parcamp pour se lier à l’opéron lac et faciliter la liaison de l’ARN polymérase au promoteur pour transcrire les gènes pour l’utilisation du lactose.
En tant que forme de répression des catabolites, l’effet glucose sert une fonction utile chez les bactéries: il oblige les cellules à utiliser la meilleure source d’énergie disponible. Pour de nombreuses bactéries, le glucose est le plus commun et le plus prêtutilisésubstrat pour la croissance. Ainsi, il inhibe indirectement la synthèse des enzymes qui métabolisent les sources d’énergie les plus pauvres.
FIN DU CHAPITRE
Page précédente
Retour à la Page 1