
okasolusyöpä (SCC), joka tunnetaan myös piikkisolusyöpänä, ovat maailmanlaajuisesti yleisimpiä syöpäkuolemien aiheuttajia. SCC on pahanlaatuinen epiteelikasvain, joka syntyy kudoksissa, jotka tarjoavat esteen organismin ja ympäristön, kuten ihon, suun, ontelon, ruokatorven ja keuhkojen välillä . Varhaisemmassa vaiheessa SCC: hen liittyy epidermaalinen keratinisaatio ja haavaumien muodostuminen limakalvon pintaan, ja myöhemmässä vaiheessa SCCs tunkeutuu syvempiin kudoksiin. Mikroskooppista esiintymistä varten SCC-solut ovat järjestäytyneet pesiin, joita ympäröivät monikulmaiset solut, joilla on erilliset solurajat ja hyperkromaattiset tumat. SCCs on korkea taipumus metastasizeed, yleensä alueellisten imusolmukkeiden kautta, ja saattaa aiheuttaa järjestelmällisiä vaurioita useita elimiä. Leikkaus, laserhoito, ja säteily ovat edelleen yleisimmin käytetty hoito SCCs . SCC-varianttien ainutlaatuiset sytomorfologiset piirteet johtavat erillisiin hoitoihin ja lopputuloksiin .
solun pinta-antigeeni CD109 on noin 170 kDa: n glykosyylifosfatidyylinositoliin (GPI) sidottu glykoproteiini ja kuuluu tioesteripitoisten proteiinien A2 makroglobuliini (a2M) / C3, C4, C5-perheeseen . Ihmisen CD109-geeni sijaitsee kromosomissa 6q ja muodostaa noin 3,3% koko CD109-genomisarjasta 33 eksoninsa kanssa . Eristetty CD109 cDNA sisältää 4335 bp: n avoimen lukukehyksen, joka koodaa 1445 aminohappoa (aa) . CD109-proteiini sisältää 21 aa n-terminaalisen leader–peptidin, oletetun syöttialueen (aa 651-683), tioesterisitoutumiskohdan (aa 918-924), heksapeptidin määrittelevän tioesterireaktiivisuuden (aa1030-1035) ja C-terminaalisen konsensus-GPI-ankkurisignaalijakson, jonka pilkkoutumisen ennustetaan tapahtuvan aminohappo 1420: n jälkeen (kuva. 1) . N-terminaalinen leader-peptidi ankkuroi proteiinin sisäkalvoon periplasmaan ja N-terminaaliseen cd109-fragmenttiin, joka erittyy soluista furiiniproteaasin pilkkouduttua . Pitkälti häiriintyneen syöttialueen proteaasi-pilkkoutuminen aktivoi CD109: n, mikä johtaa konformaatiomuutokseen, joka vangitsee proteaasin häkkimäiseen rakenteeseen ja altistaa erittäin reaktiivisen tioesterisidoksen. Tioesterisitoutumiskohta, joka aktivoituessaan voi kovalenttisesti yhdistää CD109: n hyökkäävän proteaasin pinnalla oleviin lysiinijäämiin. Lisäksi C-terminaalinen konsensus GPI-ankkuri-signaalijakso tunnetaan myös reseptoreita sitovana domeenina .

The structure of CD109 protein. CD109-proteiinin kokonaispituus koostuu 1445 aminohaposta (aa), mukaan lukien 21 aa n–terminaalinen signaalipeptidi, todennäköinen syöttialue (aa 651-683), tioesterisitoutumiskohta (aa 918-924), heksapeptidin määrittelevä tioesterireaktiivisuus (aa1030-1035) ja C-terminaalinen konsensus GPI-ankkurisignaalijakso (aa 1420-1445), jonka pilkkoutumisen arvioidaan tapahtuvan aminohapon jälkeen .1420
cd109 tunnistettiin ensin solupinta-antigeeniksi monoklonaalisella vasta-aineella, joka nostettiin primitiivistä lymfoidi/myeloidista solulinjaa kg1a vastaan. Aiemmissa tutkimuksissa on raportoitu, että CD109 ilmaistaan sikiön ja aikuisen CD34+ – positiivisten luuytimen mononukleaarisolujen, aktivoitujen t-lymfoblastien, aktivoitujen verihiutaleiden, endoteelisolujen , mesenkymaalisten kantasolujen ja useiden ihmisen kasvainsolulinjojen alajoukolla, mutta ei lepäävinä ihmisen T-soluina, verihiutaleina tai perifeerisen veren valkosoluina . Cd109: ää on havaittu SCCs: ssä erityisesti useista elimistä, kuten keuhkoista , ruokatorvesta , kohdun kohdunkaulasta , ihosta , peniksestä , suuontelosta ja sappirakosta . CD109 on transformoivan kasvutekijä-β1 (TGF-β1)-reseptorijärjestelmän komponentti ja säätelee negatiivisesti TGF-β1-signalointia . Funktiotutkimukset viittasivat siihen, että CD109 saattaa olla molekyylitason kohde uusien terapeuttisten lääkeaineiden kehittämisessä eri kudosperäisiä SCCs: iä varten . Tässä katsauksessa esitämme yhteenvedon CD109: n tämänhetkisestä tietämyksestä ja sen suhteesta SCCs: ään.
CD109 ekspressio SCCs: ssä
kasvainkudoksissa CD109 todettiin immunohistokemiallisesti SCCs: ssä sekä uroteelikarsinoomissa, pahanlaatuisissa melanoomissa , tyvitumaisten rintojen karsinoomissa , myksofibrosarkoomassa , epiteelisarkoomissa ja glioomassa . Erityisesti Shiraki et al. raportoidut cd109-positiiviset perivaskulaariset kasvainsolut ihmisen alemman luokan glioomakudoksissa ja hiirimallissa kertautuivat ihmisen glioomaan, mikä viittaa CD109: n keskeiseen rooliin tässä sairaudessa. Aiemmat tutkimukset ovat osoittaneet, että Cd109: n voimakas ilmentyminen SCCs: ssä ja rajoitettu ilmentyminen normaaleissa okasoluissa (Taulukko 1) . Lisäksi CD109 on hyvin ilmaistu hyvin eriytetty SCCs pikemminkin kuin kohtalaisen tai huonosti eriytetty SCCs, joten ilmaisutaso CD109 on kääntäen korreloi kasvain Luokka .
CD109 ja keuhkojen SCC
keuhkojen okasolusyöpä (LSCC) on yksi ei-pienten-cell keuhkosyöpä (NSCLC), ja osuus on noin 20-30% tapauksista nsclcs . CD109: n ekspressiota tutkittiin ihmisen keuhkosolukarsinoomissa kvantitatiivisella RT-PCR-menetelmällä, jossa CD109: n ilmentyminen oli huomattavasti suurempi levyepiteelikarsinoomissa, mutta ei adenokarsinoomissa, suurisolukarsinoomissa eikä pienisolukarsinoomissa . Lisäksi Sato et al. käytti CD109-vasta-ainetta analysoidakseen CD109: n ilmentymistä normaaleissa keuhko-ja keuhkosyöpäkudoksissa. Normaaleissa keuhkokudoksissa CD109: n ilmentyminen rajoittui lähinnä keuhkoputken ja keuhkoputken epiteelin tyvisoluihin . Keuhkosyöpäkudoksissa voimakas immunoreaktiivisuus havaittiin ensisijaisesti LSCCs: ssä, mutta ei muunlaisissa keuhkosyövissä . Nämä havainnot osoittavat, että Cd109: ää säännellään erityisesti LSCCs: ssä.
CD109 ja ruokatorven SCC
ruokatorven okasolusyöpä (ESCC) muodostavat kuudesosan kaikesta syöpään liittyvästä kuolleisuudesta, jota esiintyy enemmän Aasian maissa . ESCC: n ennuste on edelleen huono, ja ESCC: N 5-vuotinen eloonjäämisaste on noin 14-22% . Viimeaikaiset tutkimukset viittaavat siihen, että TGF-β-reseptorien muuttunut ilmentyminen edistää ESCC: n etenemistä, ja TGF-β-signaloinnin inhibitoristen proteiinien lisääntynyt ilmentyminen korreloi ESCC: n huonoon ennusteeseen . Hashimoto ym. tutkittiin cd109: n ilmentymistä ESCC: n kudoksissa kvantitatiivisella RT-PCR: llä, ja CD109: n mRNA-transkriptiota säädeltiin enemmän noin 50%: ssa Escc: stä. Myöhemmin CD109: n proteiinipitoisuutta ESCC: ssä tutkittiin immunohistokemialla kudosmikrorayseilla (TMA) . Ei-sairaassa ruokatorven kudoksessa CD109: n ilmentyminen rajoittui kerrostuneiden epiteelisolujen sytosolissa heikolla tasolla. Kaikissa TMAS: n ESCC-näytteissä pahanlaatuisten levyepiteelisolujen säikeissä näkyi voimakas CD109-värjäytyminen sytosolissa. Muissa solutyypeissä ei havaittu CD109-positiivisia värjäytymiä. Lisäksi CD109: n lauseke oli hyvin ja kohtalaisesti eriytyneissä Escc: issä korkeampi kuin heikosti erilaistuneissa .
CD109 ja kohdun kohdunkaula SCC
kohdunkaulan syöpä aiheuttaa noin 12% naisten syöpään liittyvistä kuolemista . SCCs: n osuus kaikista kohdunkaulan syövistä on 80-85 prosenttia . Zhang ym. osoitti, että cd109: n ilmentyminen oli merkitsevästi suurempi kohdunkaulan SCCs: ssä kuin endometriumin adenokarsinoomissa. He tutkivat myös CD109: n ilmentymistä viidessä ihmisen kohdunkaulan karsinooman solulinjassa ja havaitsivat cd109: n ilmentymisen korkeita tasoja kahdessa SCC: n solulinjassa.
CD109 ja ihon SCC
ihon okasolusyöpä (CSCC) on toiseksi yleisin ei-melanoomainen ihosyöpä, jonka esiintyvyys lisääntyy jatkuvasti . Vaikka CSCC on yleensä suotuisa ennuste, on vielä noin 1,5–2% potilaista kuolee tähän tautiin . Cd109: n ilmentymistä CSCCs: ssä on tutkittu tmas: n immunohistokemialla . Normaaleissa ihoissa CD109: ää esiintyi heikosti epidermaalisten solujen tyvikerroksessa, kun taas pahanlaatuisten levyepiteelisolujen säikeissä näkyi voimakasta CD109-värjäytymistä. Lisäksi CD109-lauseke korreloi kääntäen CSCC-arvosanojen kanssa. Cd109: n ilmentymä oli Escc: n tavoin korkeampi hyvin ja kohtalaisesti eriytyneissä CSC: issä kuin huonosti eriytyneissä . Peniksen okasolusyöpä (PCCC) on Cscc: n alatyyppi ja sitä on tutkittu harvoin . Dong ym. on osoitettu, että CD109-proteiinia esiintyy runsaasti PSC-solujen pahanlaatuisissa levyepiteelisoluissa verrattuna tmas-potilaiden normaaleihin peniksen kudoksiin. Lisäksi lausekekuvio validoidaan tuoreissa kirurgisissa PCC-näytteissä immunofluoresenssilla, qRT-PCR: llä ja western blottingilla, mikä viittaa siihen, että CD109 voi olla PCC: n biomarkkeri .
CD109 ja suuontelon SCC
suun okasolusyöpä (OSCC) muodostavat 2-3% kaikista syövistä maailmanlaajuisesti . Funktion häviäminen mutaatio TGF-β tyypin II reseptorin on usein tapahtuma suuontelon SCC . CD109: n ilmentymistä normaaleissa suun kudoksissa ja OSCC-kudoksissa 124 potilaalta tutkittiin immunohistokemiallisella värjäyksellä. Suuria CD109-pitoisuuksia todettiin usein SCCs: ssä ja suuontelon preemalignantissa leesioissa, mutta ei normaalissa levyepiteelissä . Lisäksi cd109: n lauseketaso oli selvästi korkeampi hyvin erilaistuneissa Oscc: issä kuin kohtalaisen tai huonosti erilaistuneissa Oscc: issä, mikä viittaa siihen, että CD109: n lauseke korreloi Oscc: n eriytymisvaiheiden kanssa . Lisäksi OSCC-solulinjat yli-ilmentävät CD109: n kiihtynyttä solujen kasvua in vitro , mikä viittaa siihen, että CD109 liittyy Oscc-solujen etenemiseen.
CD109 ja sappirakon SCC
sappirakon syöpä (GBC) on sappisyövistä aggressiivisin, ja elinajan mediaani on lyhin . Sappirakon okasolusyöpä (GSCC) osuus 3% pahanlaatuinen kasvain tämän elimen . Gbcs-kudosten alatyypit, mukaan lukien adenokarsinooma (AC), okasolusyöpä (SCC) ja adenosquamous carcinoma (ASC), tutkittiin tmas: llä immunohistokemiallisella värjäyksellä CD109-vasta-aineella. CD109-värjäys oli negatiivinen kaikissa normaaleissa sappirakon kudoksissa ja AC-kudoksissa. Samaan aikaan CD109-positiivisia soluja löytyi 86,7%: sta SCCs: stä ja 91,7%: sta ASCs: stä. Koska CD109 on selvästi ilmaistu pahanlaatuisia sqamous soluja sappirakon, CD109 voi olla diagnostinen markkeri sappirakon SCCs ja ASCs .
TGF-β-signalointireitti ja SCCs
TGF-β-signalointireitti osallistuu moniin soluprosesseihin, kuten solujen kasvuun, solujen erilaistumiseen apoptoosiin ja solujen homeostaasiin . TGF-β-ligandien perhe, TGF-β1, TGF-β2 ja TGF-β3 , sitoutuu spesifisiin transmembraanityypin I ja tyypin II seriini/treoniinikinaasireseptoreihin (TGF-ßR1 ja TGF-ßR2), jolloin TGF-ßR1-kinaasiaktiivisuus aktivoituu . Tämän jälkeen aktivoitu TGF-ßR1 levittää signaalia fosforyloimalla solunsisäisiä substraattejaan, R-Smadeja (Smad2 ja Smad3) . Smad2 ja Smad3 vuorovaikuttavat TGF-ßR1: n ja Saran (Smad anchor for receptor activation) kanssa, joka on Fyve-domeenin proteiini, joka vuorovaikuttaa suoraan Smad2: n ja Smad3: n kanssa , SARA toimii värvätäkseen Smad2: n TGF-β-reseptoriin, minkä jälkeen fosforyloidut R-Smadit muodostavat heteromeerisiä komplekseja Co-SMAD: n (Smad4) kanssa . Fosforylaation ja sitä seuranneiden smad4-kompleksien jälkeen nämä R-Smads-kompleksit vapautuvat TGF-ßR1: stä ja Sarasta ja translokoituvat sitten tumaan , jossa ne vuorovaikuttavat transkriptiotekijöiden kanssa, jotka värväävät ne kohdegeenien tiettyihin promoottorielementteihin .
reseptorin endosytoosi on keskeinen säätelymekanismi signaalien transduktiossa. TGF-β-reseptorit internalisoituvat sekä klatriini – että caveolae-riippuvaisten reittien kautta. TGF-β-reseptorien sisäistäminen klatriinipäällysteisten kuoppien kautta on yhdistetty signalointiin Smad2/3: n ja reseptorien kierrätyksen kautta. Sen sijaan TGF-β-reseptorin lokalisointi luolissa liittyy smad2/3-signaloinnin alasäätelyyn ja reseptorin hajoamiseen E3-ubikitiiniligaasi Smurf2: n ubikitinaation jälkeen . Kuitenkin inhibitory Smads (Smad6 ja Smad7) muodostavat erillisen alaluokan Smads jotka toimivat vastakkaisella tavalla R-Smads ja antagonize signalointi . Ne voivat kilpailla R-Smadien kanssa sitoutumisesta aktivoituneeseen TGF-ßR1: een ja siten estää R-Smadien fosforylaation . Lisäksi ne värväävät aktivoituun TGF-ßR1: een E3-ubikitiiniligaaseja, mikä johtaa reseptorin ubikitinaatioon ja hajoamiseen sekä signaloinnin päättymiseen .
TGF-β-reitin säätelyhäiriö on yhdistetty useisiin syöpätyyppeihin . Tutkimukset ovat osoittaneet, että TGF-β-signalointi saa aikaan ennaltaehkäisevän vaikutuksen tumorigeneesin varhaisemmissa vaiheissa, mutta tukahduttavan vaikutuksen kasvaimen myöhemmän kehityksen aikana . TGF-ßR1-geenin mutaatioita on löydetty myös ihon SCCs: stä, mikä viittaa siihen, että TGF-β: n inaktivoituminen johtaa SCCs: n aloittamiseen .
CD109 ja TGF-β-signalointireitti
CD109 on TGF-β-ko-reseptori ja moduloi TGF-β-signalointireseptorin toimintaa solupesifisesti . Solun pinnalla CD109 säätelee negatiivisesti TGF-β1-signalointireittiä muodostamalla reseptorikompleksin TGF-ßR1: n ja TGF-ßR2: n kanssa ihmisen keratinosyyteissä . TGF-β-reseptorit hajoavat E3-ubikitiiniligaasi Smurf2: n ubikitinaation jälkeen ja internalisoituvat sekä klatriiniriippuvaisten että luolaliippuvaisten reittien kautta . Bizet ym. osoitettu, että CD109 liittyy caveolin-1: een ja edistää TGF-β-reseptorin endosytoosia. Lisäksi CD109 edistää TGF-β-reseptorien lokalisointia kaveolaariseen lokeroon ligandin läsnä ollessa ja helpottaa TGF-β-reseptorin hajoamista. CD109 säätelee myös smad7/Smurf2: n lokalisointia ja liittymistä TGF-ßR1: een. CD109: n estovaikutukset vaativat Smad7: n ilmentymistä ja Smurf2: n ubikitiiniligaasin aktiivisuutta . Lisäksi CD109: ää voi vapautua solun pinnalta solulipaasien kuten fosfatidyylinositolispesifisen fosfolipaasi C: n (PI-PLC) avulla. Cd109: n liukoinen muoto säilyttää kykynsä sitoa TGF-β1 ja takavarikoida se pois TGF-β-reseptoreista .
kuitenkin Vorstenbosch et al. raportoitu, että CD109 eri säännelty TGF-β-indusoitu ALK1-Smad1 / 5 vs. ALK5-Smad2/3 polkuja (ALK1 ja ALK5 ovat kaikki TGF-β tyypin I reseptoreihin). He havaitsivat, että TGF-β-signalointi estää endoteelisolujen proliferaatiota ja migraatiota, kun taas TGF-β-signalointi indusoi näitä prosesseja myös alk1-Smad1/5: n kautta . Ne osoittavat, että ALK1 ilmaistaan ja rinnakkaispaikallistuu cd109: n kanssa hiiren keratinosyyteissä ja että hiiret yliekspressoivat CD109: ää orvaskedessä näyttävät tehostettua ALK1-Smad1/5-signalointia, mutta vähenivät ALK5-Smad2/3-signalointia .
lisäksi TGF-β1 on voimakas kasvun estäjä useimmissa epiteelisoluissa . Hagiwara ym. osoitettu, että oraaliset SCC-solulinjat yli-ekspressio CD109 nopeutti solujen proliferaatiota ja heikensi TGF-β1: n välittämää antiproliferatiivista vaikutusta. Sen sijaan SCC-soluissa, joissa oli cd109: n knockdown, solujen kasvu oli hitaampaa . Cd109: n voimakas ilmentyminen esti Smad2-fosforylaatiota, mikä heikensi TGF-β1/Smad2-signalointia ja heikentää TGF-β1-välitteistä solujen kasvun estoa, CD109 knockdown lisäsi Smad2-fosforylaatiota TGF-β1-stimulaation avulla . Vaikka CD109 säätelee myös Smad1 / 5-signalointia , sitä ei ole liitetty SCC: n kehitykseen. Yhdessä CD109 helpottaa SCCs: n kehittymistä estämällä TGF-β-Smad2/3-reittiä (Kuva. 2).

CD109 ja TGF-β-signalointi. TGF-β sitoo TGF-ßR2: n, joka värvää ja fosforyloi TGF-ßR1: n. Aktivoitu TGF-ßR1 levittää signaalia myötävirtaan fosforyloimalla suoraan Smad2: ta ja Smad3: a. Fosforyloidut R-Smadit muodostavat tämän jälkeen heteromeerisiä komplekseja smad4: n kanssa yhdessä transkriptiotekijöiden kanssa, säätelevät geenien transkriptiota ja solujen toimintaa. TGF-β-reseptorit internalisoituvat klatriinipäällysteisten kuoppien tai kaveolaarisen reitin kautta. CD109 lisää TGF-β: n sitoutumista TGF-β-reseptoreihin ja edistää TGF-β-reseptorin lokalisointia luoliin, lisää lopulta TGF-β-reseptorin endosytoosia ja helpottaa TGF-β-reseptorin hajoamista. CD109: n estovaikutukset edellyttävät Smad7: n ilmentymistä ja Smurf2 ubikitiiniligaasin aktiivisuutta
CD109 ja STAT3-signalointi
transkriptiotekijän 3 (STAT3) signaalianturi ja aktivaattori on kriittinen solujen proliferaation, erilaistumisen, migraation, eloonjäämisen ja onkogeneesin kannalta . Litvinov ym. raportoitu, että cd109-proteiinin ekspressio oli selvästi vähentynyt psoriaattisessa orvaskedessä verrattuna viereiseen epähygieeniseen ihoon. Cd109 mRNA-ekspressio on kuitenkin psoriasisplakeilla muuttumaton verrattuna normaaliin ihoon, mikä viittaa siihen, että cd109-proteiinin vapautuminen lisääntyy psoriaattisissa keratinosyyteissä . He ehdottivat, että vapautunut/liukeneva CD109 pystyy indusoimaan molekyylimuutoksia, joita tiedetään esiintyvän psoriaasissa . In vitro, he havaitsivat, että transfektio CD109 siRNA alas säätelee STAT3, vapauttaa CD109 solujen pinnalta viljeltyjen ihmisen keratinosyyttien. Lisäksi eksogeeninen / rekombinantti CD109 indusoi STAT3-signalointia ihmisen keratinosyyteissä . Sitä paitsi, Chuang ym. raportoitu, että CD109 expression oli dramaattinen regulated metastaattinen keuhkojen adenokarsinooma soluja, ja solut ilmentävät CD109 shRNA (shCD109) osoitti dramaattinen väheneminen STAT3 fosforylaatio. STAT3 knockdown vähensi huomattavasti etäpesäkkeitä, ja STAT3: n aktiivisuuden palauttaminen lisäsi shCD109: ää ilmentävien solujen kykyä etäpesäkkeisiin . Aktivaation yhteydessä STAT3 fosforyloituu ei-reseptoriproteiini tyrosiinikinaasien janus-kinaasi 2: n (JAK2) vaikutuksesta, jolloin muodostuu STAT3-dimeeri ja translokaatio tumaan . Jak-kinaasiaktiivisuuden esto fibroblastien yli-ilmentävässä cd109: ssä kuitenkin vähensi FOSFORYLOITUNEEN STAT3: n tasolle, joka on sama kuin vanhempien soluissa, jotka ilmentävät alhaisia CD109-pitoisuuksia, mikä viittaa siihen, että CD109: n aiheuttama STAT3-fosforylaatio vaatii JAK-kinaasiaktiivisuutta. Siten JAK / STAT3-signalointi saattaa välittää CD109: n vaikutuksia kasvaimen kasvuun ja etäpesäkkeisiin .
vaikka CD109: n tyrmäys ihmisen keratinosyyteissä ja keuhkojen adenokarsinoomasoluissa vähensi STAT3-signalointia in vitro , CD109-puutoksesta kärsineillä hiirillä tulokset olivat päinvastaiset. Mii ym. cd109-vajavaiset hiiret, joilla esiintyi ihopoikkeavuuksia, mukaan lukien epiteelin hyperplasia ja tulehdussolujen infiltraatio. He raportoivat, että STAT3-fosforylaatio CD109-puutteellisilla hiirillä oli huomattavasti suurempi verrattuna villityyppisiin hiiriin. Lisäksi STAT3-signaloinnin säätely liittyy lisääntyneeseen proliferaatioon ja keratinosyyttien heikentyneeseen erilaistumiseen .
in vitro-ja in vivo-tutkimusten tulosten ero saattaa johtua cd109-puutoksesta kärsivien hiirten kudoksissa tapahtuneista mikroympäristön systemaattisista muutoksista. CD109: n häviäminen kaikista hiirten soluista saattaa muuttaa ihonalaista mikroympäristöä, joka aktivoi STAT3-signalointia keratinosyyteissä. Lisäksi CD109: llä voi olla erillisiä säätelyvaikutuksia eri solutyypeissä, mikä johtaa solutyyppikohtaisiin muutoksiin STAT3-signaloinnissa. Keratinosyyttien lisäksi CD109: ää esiintyy endoteelisoluissa, epiteelisoluissa ja fibroblasteissa, jotka osallistuvat ihokudoksen muodostamiseen. CD109: n ja STAT3-signaloinnin suhdetta ei kuitenkaan ole toistaiseksi tutkittu näissä solutyypeissä.
CD109 ja EGFR-signalointi
epidermaalinen Kasvutekijäreseptori (EGFR) kuuluu erbb-reseptoriperheeseen. EGF: n sitoutuessa ligandiin EGFR muodostaa dimeerejä, joko homodimeerejä tai heterodimeerejä toisen erbb-suvun jäsenen HER2: n kanssa . Dimerisoituneet reseptorit auto-fosforyloivat toisensa ja fosforyloivat sitten ei-reseptoriproteiini tyrosiini C-Src-kinaasin, joka aktivoi STAT3: n . EGFR: n aktivointi edistää solujen siirtymistä, eloonjäämistä ja proliferaatiota. Pahanlaatuisissa kasvaimissa EGFR-yliilmaisu korreloi kasvaimen invaasion syvyyden kanssa ja liittyy huonompaan ennusteeseen . EGFR: n yli-ekspressioon johtavia mutaatioita havaitaan keuhkojen SCC: ssä , pään ja kaulan SCC: ssä sekä ruokatorven SCC: ssä . SK-MG-1-soluissa oleva kalvoankkuroitu CD109 on suoraan vuorovaikutuksessa EGFR: n kanssa ja tehostaa EGF-signalointia, mikä myöhemmin lisää solujen muuttoliikettä ja invaasiota, kun taas erittyvä CD109 ei vaikuta EGF-signalointiin . EGFR saattaa välittää CD109: n vaikutuksia STAT3-signalointiin, mikä vaatii lisätutkimuksia selventääkseen (Kuva. 3).
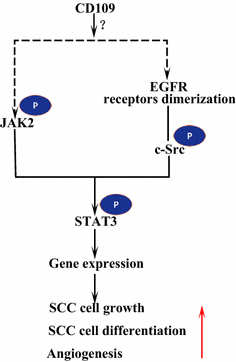
CD109 ja EGFR / STAT3 signalointi. CD109 helpottaa EGFR-signalointia. Dimerisoituneet EGFR-reseptorit fosforyloivat c-Src: tä, joka puolestaan fosforyloi STAT3: a. CD109 voi myös aktivoida STAT3: n JAK2: n fosforylaation kautta, minkä jälkeen se edistää SCC-solujen kasvua, erilaistumista ja angiogeneesiä
tähän mennessä tumorigeeneesin ja SCC: n kehittymisen kokonaiskuva on edelleen epätäydellinen. TGF-β: n ja STAT3: n tai EGFR: n signalointireittien välillä on ristikeskusteluja. CD109 on todennäköisesti yksi SCCs: ää säätelevän signalointiverkon tärkeimmistä efektoreista. Cd109: n roolista SCCs: n patogeneesissä ei kuitenkaan ole raportoitu suoria todisteita.
Perspectives
ihmisen kudosnäytteistä tehdyt tutkimukset osoittavat, että CD109: ää esiintyy voimakkaasti useiden elinten SCCs: ssä, erityisesti hyvin erilaistuneissa pahanlaatuisissa okasoluissa . Osoittamalla CD109: n ilmentymistä immunohistokemialla ihmiskudoksissa CD109 voi mahdollisesti toimia biomarkkerina SCC: n etenemisen määrittämiseksi. Nykyisten tutkimusten mukaan CD109 ilmaistaan hyvin erilaistuneissa SCCs: issä ja sen ilmentyminen on vähäisempää erilaistumattomissa SCCs: issä . Ei kuitenkaan ole selvää, liittyykö CD109 verisuonten invaasioon, etäpesäkkeisiin ja ennusteeseen leikkauksen jälkeen. Siksi tarvitaan lisätutkimuksia CD109: n kliinis-patologisen merkityksen selvittämiseksi SCCs: ssä suuremmassa otoskoossa.
CD109 on GPI: hen liittyvä glykoproteiini, jonka avulla sitä voidaan vapauttaa kalvosta . Cd109: n liukoinen muoto vaikuttaa myös TGF-β: n sitoutumiseen reseptoreihin ja moduloi SCC: n etenemistä . Litvinov ym. todettiin, että CD109 vapautuu solun pinnalta solunulkoiseen ympäristöön, ja cd109: n vapautunut muoto säilyttää kykynsä indusoida solunsisäisiä signalointireittejä. Sitä paitsi, Sakakura et al. raportoitu, että seerumin CD109 julkaistiin xenografted kasvain ja se kasvaa suhteessa määrän kasvaimen xenograft. Siksi seerumin CD109-tason havaitseminen saattaa auttaa valvomaan yli-ilmentyviä kasvaimia CD109 mukaan lukien SCCs.
Eksosomit viestivät primaarisesta kasvainleesiosta ja sen lokerosta valikoituja proteiineja tai muita molekyylejä sisältävän paketin kautta . Kiertävän kasvaimeen liittyvän eksosomin eristäminen ja analysointi voivat toimia biomarkkereina syöpäpotilaiden diagnosoinnissa. Eksogeeninen CD109 on tunnistettu transfektoiduista 293 solusta erittyvän eksosomin komponentiksi, joten se on lupaava kohde eksosomin diagnosoinnissa . Tähän mennessä CD109: ää ei ole kuitenkaan raportoitu SCC: stä tai muista kasvainsoluista peräisin olevissa eksosomeissa. CD109: n exosomeihin pakkaamisen taustalla olevia mekanismeja on syytä tutkia tarkemmin.
biomarkkerin potentiaalin lisäksi CD109 saattaa olla terapeuttisten lähestymistapojen kohde. CD109 on kalvoproteiini, johon voidaan kohdistaa suoraan spesifisiä vasta-aineita tai entsyymejä. CD109 saatetaan tunnistaa myös huumeiden jakeluun tarkoitetuista kohdennetuista ajoneuvoista. CD109: n yksityiskohtaiset roolit SCCs: n patogeneesissä ovat kuitenkin vielä epäselviä. Esimerkiksi cd109: n ja erilaistumattoman SCCs: n alemman lausekkeen suhde on määriteltävä. CD109-interventiota voidaan harkita kliinisessä käytännössä vasta, kun sen riskit ja hyödyt on arvioitu huolellisesti.