El carcinoma de células escamosas (CCE), también conocido como carcinoma de células espinosas, es una de las principales causas de muerte por cáncer en todo el mundo. El CCE es un tumor epitelial maligno , que surge en tejidos que proporcionan una barrera entre un organismo y el medio ambiente, como la piel, la boca, la cavidad, el esófago y el pulmón . En una etapa temprana, el CCE se acompaña de queratinización epidérmica y formación de úlceras en la superficie de la mucosa, y los tejidos más profundos son invadidos por los CCE en una etapa posterior. Para la apariencia microscópica, las células CCE están dispuestas en nidos, que están rodeados de células poligonales con bordes celulares distintos y núcleos hipercromáticos. Los CCE tienen una alta tendencia a hacer metástasis, por lo general a través de los ganglios linfáticos regionales, y pueden causar daños sistemáticos en múltiples órganos. La cirugía, la terapia con láser y la radiación siguen siendo los tratamientos más utilizados para los CCE . Las características citomorfológicas únicas de las variantes de CCE conducen a tratamientos y resultados distintos .
El antígeno de superficie celular CD109 es una glicoproteína ligada al glicosilfosfatidilinositol (GPI) de aproximadamente 170 kDa y un miembro de la familia de proteínas que contienen tioéster a2 macroglobulina (a2M)/C3, C4 y C5 . El gen CD109 humano se encuentra en el cromosoma 6q, constituyendo aproximadamente el 3,3% de la secuencia genómica CD109 total con sus 33 exones . El ADNc CD109 aislado comprende un marco de lectura abierta de 4335 bp que codifica un aminoácido 1445 (aa). La proteína CD109 contiene un péptido líder N-terminal 21 aa, una región de cebo putativo (aa 651-683), un sitio de unión de tioéster (aa 918-924), un hexapéptido definidor de reactividad de tioéster (aa1030–1035) y una secuencia de señal de anclaje GPI de consenso C-terminal con la escisión prevista para ocurrir después del aminoácido 1420 (Fig. 1) . El péptido líder N-terminal ancla la proteína a la membrana interna dentro del periplasma y el fragmento N-terminal de CD109 secretado de las células después de la escisión por la proteasa de furina . La escisión de proteasa de una región de cebo en gran parte desordenada activa CD109, lo que resulta en un cambio conformacional que atrapa a la proteasa en una estructura similar a una jaula y expone el enlace tioéster altamente reactivo. El punto de unión del tioéster que, al activarse, puede enlazar covalentemente CD109 con residuos de lisina en la superficie de la proteasa atacante. Además, la secuencia de señal de anclaje GPI de consenso C-terminal también se conoce como dominio de unión al receptor .
Fig. 1
The structure of CD109 protein. La longitud total de la proteína CD109 comprende 1445 aminoácidos (aa), incluido un péptido de señal N-terminal de 21 aa, una región de cebo putativa (aa 651-683), un sitio de unión de tioéster (aa 918-924), un hexapéptido que define la reactividad de tioéster (aa1030-1035) y una secuencia de señal de anclaje GPI de consenso C–terminal (aa 1420-1445) con la escisión que se predice que ocurrirá después del aminoácido 1420
El CD109 se identificó por primera vez como antígeno de superficie celular por un anticuerpo monoclonal elevado contra la línea celular linfoide/mieloide primitiva KG1a . Estudios previos informaron que CD109 se expresa en un subconjunto de células mononucleares de médula ósea positivas para CD34+, linfoblastos T activados, plaquetas activadas, células endoteliales, subconjuntos de células madre mesenquimales y varias líneas celulares tumorales humanas , pero no se expresa en células T humanas en reposo, plaquetas o leucocitos de sangre periférica . En particular, se ha detectado CD109 en CCE de varios órganos, incluidos pulmón , esófago , cuello uterino , piel , pene , cavidad oral y vesícula biliar . CD109 es un componente del sistema receptor del factor de crecimiento transformador-β1 (TGF-β1) y regula negativamente la señalización del TGF-β1 . Los estudios de pérdida y ganancia de funciones sugirieron que CD109 podría ser un objetivo molecular para el desarrollo de nuevas terapias para CCE de diversos orígenes tisulares . En esta revisión, presentaremos un resumen de los conocimientos actuales sobre CD109 y su relación con los CCSC.
Expresión de CD109 en CCE
En tejidos tumorales, el CD109 se detectó inmunohistoquímicamente en CCE , así como en carcinomas uroteliales , melanomas malignos , carcinomas de mama de tipo basal , mixofibrosarcoma, sarcomas epiteliales y glioma . En particular, Shiraki et al. se notificaron células tumorales perivasculares CD109 positivas en tejidos de glioma humano de grado inferior y en un modelo de ratón de glioma humano recapitulado, lo que indica una función clave de CD109 para esta enfermedad. Estudios previos han demostrado que la alta expresión de CD109 en CCE y la expresión limitada en células escamosas normales (Tabla 1) . Además, el CD109 se expresa en gran medida en CCE bien diferenciados en lugar de en CCE moderadamente o pobremente diferenciados, por lo que el nivel de expresión de CD109 se correlaciona inversamente con el grado tumoral .
Tabla 1 Expresión de CD109 en carcinomas de células escamosas
CD109 y CCE de pulmón
El carcinoma de células escamosas de pulmón (CSCP) es uno de los subtipos principales de cáncer de pulmón de células pequeñas (CPCNP), y representa aproximadamente 20-30% de los casos de CPCNP . Se examinó la expresión de CD109 en carcinomas de células pulmonares humanas mediante RT-PCR cuantitativa, que mostró una expresión significativamente más alta de CD109 en carcinomas de células escamosas, pero no en adenocarcinomas, carcinomas de células grandes o carcinomas de células pequeñas . Además, Sato et al. se utilizó un anticuerpo CD109 para analizar la expresión de CD109 en tejidos normales de pulmón y carcinoma de pulmón. En los tejidos pulmonares normales, la expresión de CD109 se limitó principalmente a las células basales del epitelio bronquial y bronquiolar . En los tejidos del carcinoma de pulmón, la inmunorreactividad fuerte se detectó preferentemente en los CCL, pero no en otros tipos de carcinomas de pulmón . Estos hallazgos indican que el CD109 está específicamente regulado en los CCLL.
CD109 y CCE de esófago
El carcinoma de células escamosas de esófago (CCE) representa una sexta parte de la mortalidad relacionada con el cáncer, con una incidencia más alta en los países asiáticos . El pronóstico del ESCC sigue siendo precario, y la tasa de supervivencia a 5 años de ESCC es de aproximadamente 14-22% . Estudios recientes sugieren que la expresión alterada de los receptores TGF-β contribuye a la progresión de la ESCC, y la expresión elevada de proteínas inhibidoras de la señalización de TGF-β se correlaciona con un mal pronóstico de la ESCC . Hashimoto et al. se examinó la expresión de CD109 en los tejidos del CEC mediante RT-PCR cuantitativa, y la transcripción del ARNm del CD109 se reguló al alza en aproximadamente el 50% de los CEC. Posteriormente, se examinó el nivel de proteína CD109 en ESCC mediante inmunohistoquímica en microarrays de tejidos (MAT) . En el tejido esofágico no enfermo, la expresión de CD109 se restringió en el citosol de las células epiteliales estratificadas a un nivel débil. En todas las muestras de ESCC de las ATM, las hebras de células epiteliales escamosas malignas mostraron una fuerte tinción de CD109 en el citosol. No se observó tinción CD109 positiva en otros tipos celulares. Además, la expresión de CD109 fue mayor en los CCC bien y moderadamente diferenciados que en los escasamente diferenciados .
CD109 y CCE de cuello uterino
El cáncer de cuello uterino (cuello uterino) representa aproximadamente el 12% de las muertes de mujeres relacionadas con el cáncer . Los CCE representan entre el 80 y el 85% de todos los cánceres de cuello uterino . Zhang et al. mostró que la expresión de CD109 fue significativamente más alta en los CCE cervicales que en los adenocarcinomas endometriales. También investigaron la expresión de CD109 en cinco líneas celulares de carcinoma cervical humano y observaron niveles altos de expresión de CD109 en dos líneas celulares de CCE.
CD109 y CCE cutáneo
El carcinoma cutáneo de células escamosas (CSCC) es el segundo tipo más común de cáncer de piel no melanoma (CPNM) con una incidencia en constante aumento . Aunque el CSCC tiene un pronóstico generalmente favorable, todavía hay aproximadamente 1,5–2% de pacientes que mueren por esta enfermedad . La expresión de CD109 en CCSC ha sido examinada por inmunohistoquímica en TMAs . En las pieles normales, CD109 se expresó débilmente en la capa basal de células epidérmicas, mientras que las hebras de células epidérmicas escamosas malignas mostraron una fuerte tinción de CD109. Además, la expresión CD109 se correlacionó inversamente con los grados CSCC. Al igual que los CCC, la expresión de CD109 fue mayor en los CCC bien y moderadamente diferenciados que en los pobremente diferenciados . El carcinoma de células escamosas de pene (CPSC) es un subtipo de CCSC y se ha estudiado en raras ocasiones . Dong et al. se demostró que la proteína CD109 se expresa en gran medida en las células escamosas malignas de los CPCS en comparación con los tejidos normales del pene en las MTA. Además, el patrón de expresión se valida en muestras de CPSC quirúrgicas frescas mediante inmunofluorescencia, PCR qRT y western blot, lo que sugiere que el CD109 puede ser un biomarcador para el CPSC .
El CD109 y el CCE de cavidad oral
El carcinoma de células escamosas orales (CEC) representan del 2 al 3% de todos los cánceres en todo el mundo . La pérdida de mutación funcional en el receptor TGF-β de tipo II es un evento frecuente para el CCE de la cavidad oral . La expresión de CD109 en tejidos orales normales y tejidos OSCC de 124 pacientes se examinó mediante tinción inmunohistoquímica. Se detectaron con frecuencia niveles altos de expresión de CD109 en CCE y lesiones premalignas de la cavidad oral, pero no en epitelios escamosos normales . Por otra parte, el nivel de expresión de CD109 fue significativamente mayor en OSCC bien diferenciados que en OSCC moderadamente o pobremente diferenciados, lo que implica que la expresión de CD109 está correlacionada con las etapas de diferenciación de OSCC . Además, las líneas celulares OSCC que sobreexpresan CD109 exhibieron un crecimiento celular acelerado in vitro, lo que implica que CD109 participa en la progresión de los OSCC.
CD109 y CCE de vesícula biliar
El cáncer de vesícula biliar (GBC) es el más agresivo de los cánceres biliares con mediana de supervivencia más corta . El carcinoma de células escamosas de la vesícula biliar (CCG) representa el 3% de la neoplasia maligna de este órgano . Se examinaron subtipos de tejidos de GBCs, como adenocarcinoma (AC), carcinoma de células escamosas (CCE) y carcinoma adenoescamoso (ASC) en MAT mediante tinción inmunohistoquímica con un anticuerpo CD109. La tinción CD109 fue negativa en todos los tejidos normales de la vesícula biliar y los tejidos AC. Mientras tanto, se encontraron células CD109 positivas en el 86,7% de los CCSC y el 91,7% de los CCSC. Dado que el CD109 se expresa claramente en células escamosas malignas de la vesícula biliar, el CD109 puede ser un marcador diagnóstico de los CCE y los CCA de la vesícula biliar .
Vía de señalización TGF-β y CCE
La vía de señalización TGF-β está involucrada en muchos procesos celulares, incluyendo el crecimiento celular, la apoptosis de diferenciación celular y la homeostasis celular . La familia de ligandos TGF-β, TGF-β1, TGF-β2 y TGF-β3, se une a receptores específicos de serina/treonina quinasa tipo I y tipo II de la transmembrana (TGF-ßR1 y TGF-ßR2) , lo que resulta en la activación de la actividad de la quinasa TGF-ßR1 . El TGF-ßR1 activado propaga la señal fosforilando sus sustratos intracelulares, R-SMADs (Smad2 y Smad3). Smad2 y Smad3 interactúan con TGF-ßR1 y SARA (ancla Smad para la activación del receptor), una proteína de dominio FYVE que interactúa directamente con Smad2 y Smad3 , SARA funciona para reclutar Smad2 al receptor TGF-β, luego las R-SMADs fosforiladas forman complejos heteroméricos con Co-SMAD (Smad4). Después de la fosforilación y los complejos posteriores con Smad4, estos complejos R-Smads se liberan de TGF-ßR1 y SARA , y luego se trasladan al núcleo donde interactúan con factores de transcripción que los reclutan para elementos promotores específicos de genes diana .
La endocitosis del receptor es un mecanismo regulador fundamental en la transducción de señales. Los receptores TGF-β se internalizan a través de vías dependientes de clatrina y caveola. La internalización de los receptores TGF-β a través de las fosas recubiertas de clatrina se ha relacionado con la señalización a través de Smad2/3 y el reciclaje de receptores. Por el contrario, la localización del receptor TGF-β en caveolas se asocia con la regulación a la baja de la señalización Smad2/3 y la degradación del receptor después de la ubiquitinación por la pitufa E3-ubiquitina ligasa 2 . Sin embargo, los Smads inhibitorios (Smad6 y Smad7) forman una subclase distinta de Smads que actúan de manera opuesta a los R-Smads y antagonizan la señalización . Pueden competir con los R-Smads para unirse al TGF-ßR1 activado y, por lo tanto, inhibir la fosforilación de los R-Smads . Además, reclutan ligasas E3-ubiquitina para el TGF-ßR1 activado, lo que resulta en la ubiquitinación y degradación del receptor, y la terminación de la señalización .
La desregulación de la vía TGF-β se ha relacionado con múltiples tipos de cáncer . Los estudios han demostrado que la señalización del TGF-β provoca un efecto preventivo durante las primeras etapas de la génesis tumoral, pero un efecto supresor durante el desarrollo tumoral posterior . También se han encontrado mutaciones en el gen TGF-ßR1 en los CCSC de la piel, lo que sugiere que la inactivación del TGF-β conduce al inicio de los CCSC .
CD109 y vía de señalización de TGF-β
CD109 es un co-receptor de TGF-β y modula la actividad del receptor de señalización de TGF-β de una manera específica de la célula . En la superficie celular, CD109 regula negativamente la vía de señalización del TGF-β1 a través de la formación de un complejo receptor con TGF-ßR1 y TGF-ßR2 en queratinocitos humanos . Los receptores TGF-β se degradan después de la ubiquitinación por la E3-ubiquitina ligasa Pituf2 , y se internalizan a través de vías dependientes de clatrina y caveolas . Bizet et al. demostró que CD109 se asocia con caveolina – 1 y promueve la endocitosis del receptor TGF-β. Además, CD109 promueve la localización de los receptores TGF-β en el compartimento caveolar en presencia de ligando y facilita la degradación del receptor TGF-β. CD109 también regula la localización y la asociación de Smad7 / Smurf2 con TGF-ßR1. Los efectos inhibidores de CD109 requieren la expresión de Smad7 y la actividad de la ubiquitina ligasa Pituf2 . Además, CD109 puede ser liberado de la superficie celular por lipasas celulares como la fosfolipasa C específica de fosfatidilinositol (PI-PLC). La forma soluble de CD109 conserva su capacidad de unirse al TGF-β1 y confiscarlo lejos de los receptores TGF-β .
Sin embargo, Vorstenbosch et al. se informó de que CD109 regulaba diferencialmente las vías ALK1-Smad1/5 inducidas por TGF-β frente a las vías ALK5-Smad2/3 (ALK1 y ALK5 son todos receptores de tipo I de TGF-β). Encontraron que la señalización de TGF-β inhibe la proliferación y migración de células endoteliales, mientras que la señalización de TGF-β también induce estos procesos a través de ALK1-Smad1 / 5 . Demuestran que ALK1 se expresa y co-localiza con CD109 en queratinocitos de ratón y que los ratones que sobreexpresan CD109 en la epidermis muestran una señalización mejorada de ALK1-Smad1/5, pero una señalización disminuida de ALK5-Smad2/3 .
Además, el TGF-β1 es un potente inhibidor del crecimiento en la mayoría de las células epiteliales . Hagiwara et al. se demostró que la sobreexpresión de las líneas celulares de CCE orales CD109 aceleró la proliferación celular y afectó el efecto antiproliferativo mediado por el TGF-β1. En contraste, las células de CCE con CD109 derribadas exhibieron un crecimiento celular más lento . Un alto nivel de expresión de CD109 inhibió la fosforilación de Smad2, atenuando así la señalización de TGF-β1/Smad2 y perjudicando la supresión mediada por TGF-β1 del crecimiento celular, la eliminación de CD109 aumentó la fosforilación de Smad2 por estimulación de TGF-β1 . Aunque CD109 también regula la señalización Smad1/5, no se ha conectado con el desarrollo de SCC. En conjunto, CD109 facilita el desarrollo de CCE a través de la inhibición de la vía TGF-β-Smad2/3 (Fig. 2).
Fig. 2
Señalización CD109 y TGF-β. El TGF-β se une al TGF-ßR2, que recluta y fosforila el TGF-ßR1. El TGF-ßR1 activado propaga la señal aguas abajo fosforilando directamente Smad2 y Smad3. Los R-SMAD fosforilados luego forman complejos heteroméricos con Smad4, combinados con factores de transcripción, regulan la transcripción génica y la función celular. Los receptores TGF-β se internalizan a través de las fosas recubiertas de clatrina o la vía caveolar. CD109 aumenta la unión del TGF-β a los receptores TGF-β y promueve la localización del receptor TGF-β a las caveolas, en última instancia aumenta la endocitosis del receptor TGF-β y facilita la degradación del receptor TGF-β. Los efectos inhibidores de CD109 requieren la expresión de Smad7 y la actividad de la ubiquitina ligasa Smurf2
La señalización de CD109 y STAT3
El transductor de señal y activador del factor de transcripción 3 (STAT3) es fundamental para la proliferación, diferenciación, migración, supervivencia y oncogénesis celulares . Litvinov et al. se informó de que la expresión de la proteína CD109 disminuyó notablemente en la epidermis psoriásica en comparación con la piel adyacente no involucrada. Sin embargo, la expresión de ARNm CD109 no cambia en las placas psoriásicas en comparación con la piel normal, lo que sugiere la posibilidad de que la liberación de proteína CD109 aumente en los queratinocitos psoriásicos . Sugirieron que CD109 liberado / soluble es capaz de inducir cambios moleculares que se sabe que ocurren en la psoriasis . In vitro, encontraron que la transfección de CD109 siRNA disminuye STAT3, liberación de CD109 de la superficie celular de queratinocitos humanos cultivados. Además, CD109 exógeno / recombinante induce la señalización STAT3 en queratinocitos humanos . Además, Chuang et al. se informó que la expresión de CD109 se reguló dramáticamente en las células de adenocarcinoma de pulmón metastásico, y las células que expresaban un ARN CD109 (shCD109) mostraron una reducción dramática en la fosforilación de STAT3. La eliminación de STAT3 redujo en gran medida las metástasis y la restauración de la actividad de STAT3 aumentó la capacidad de las células que expresan shCD109 para hacer metástasis . Tras la activación, STAT3 es fosforilado por la proteína no receptora tirosina quinasa janus quinasa 2 (JAK2), lo que lleva a la formación de dímero STAT3 y la translocación al núcleo . Sin embargo, la inhibición de la actividad de la cinasa JAK en fibroblastos que sobreexpresan CD109 redujo el ESTAT3 fosforilado a un nivel similar al de las células parentales que expresan bajos niveles de CD109, lo que sugiere que la fosforilación de ESTAT3 inducida por CD109 requiere actividad de la cinasa JAK. Por lo tanto, la señalización JAK / STAT3 podría mediar los efectos de CD109 en el crecimiento tumoral y la metástasis .
Aunque la caída de CD109 en queratinocitos humanos y células de adenocarcinoma de pulmón regula a la baja la señalización de STAT3 in vitro , los ratones con deficiencia de CD109 mostraron resultados opuestos. Mii et al. se generaron ratones con deficiencia de CD109, que exhibieron anomalías cutáneas como hiperplasia epitelial e infiltración de células inflamatorias. Informaron que la fosforilación de STAT3 en ratones con deficiencia de CD109 fue significativamente mayor en comparación con los ratones de tipo salvaje. Además, la regulación ascendente de la señalización STAT3 se asocia con un aumento de la proliferación y un deterioro de la diferenciación de queratinocitos .
La discrepancia de los resultados de los estudios in vitro e in vivo podría deberse a los cambios sistemáticos del microambiente en los tejidos de ratones con deficiencia de CD109. La pérdida de CD109 en todas las células de ratones podría modificar el microambiente subcutáneo que activa la señalización de STAT3 en los queratinocitos. Además, el CD109 puede ejercer efectos reguladores distintos en diferentes tipos de células, lo que lleva a una modificación específica de tipo celular en la señalización STAT3. Además de los queratinocitos, CD109 se expresa en células endoteliales, células epiteliales y fibroblastos, que participan en el tejido de la piel. Sin embargo, hasta la fecha, la relación de la señalización CD109 y STAT3 no se ha explorado en estos tipos de células.
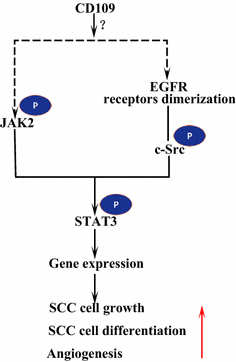
El receptor del factor de crecimiento epidérmico (EGFR) es un miembro de la familia de receptores ErbB. Tras la unión del ligando por EGF, el EGFR forma dímeros, homodímeros o heterodímeros con otro miembro de la familia ErbB HER2 . Los receptores dimerizados se fosforilan entre sí y luego fosforilan la proteína tirosina c-Src quinasa no receptora, que activa STAT3 . La activación del EGFR promueve la migración celular, la supervivencia y la proliferación. En los tumores malignos, la sobreexpresión de EGFR se correlaciona con la profundidad de invasión del tumor y se relaciona con un pronóstico más precario . Las mutaciones que conducen a la sobreexpresión de EGFR se detectan en el CCE de pulmón , el CCE de cabeza y cuello y el CCE de esófago . El CD109 anclado a membrana en las células SK-MG-1 interactúa directamente con el EGFR y mejora la señalización del EGF, lo que posteriormente aumenta la migración e invasión celular, mientras que el CD109 secretado no tiene efecto en la señalización del EGF . El EGFR podría mediar en los efectos de CD109 en la señalización de STAT3, lo que requiere más estudios para dilucidar (Fig. 3).
Fig. 3
CD109 y EGFR/STAT3 señalización. CD109 facilita la señalización EGFR. Los receptores EGFR dimerizados fosforilan c-Src, que a su vez fosforilan STAT3. CD109 también puede activar STAT3 a través de la fosforilación de JAK2, luego promover el crecimiento, la diferenciación y la angiogénesis de las células CCE
Hasta la fecha, el panorama general de la tumorogénesis y el desarrollo de CCE sigue siendo incompleto. Hay conversaciones cruzadas entre las vías de señalización TGF-β y STAT3 o EGFR. El CD109 es probablemente uno de los efectores clave de la red de señalización que regula los SCCS. Sin embargo, no se han notificado pruebas directas que definan las funciones de CD109 en la patogénesis de los CCSc.
Perspectivas
Los estudios de muestras de tejido humano indican que el CD109 está altamente expresado en CCE de múltiples órganos, en particular en células escamosas malignas bien diferenciadas . Mediante la detección de la expresión de CD109 con inmunohistoquímica en tejidos humanos, CD109 puede actuar potencialmente como un biomarcador para determinar la progresión del CCE. Los estudios actuales sugieren que CD109 está altamente expresado en CCSC bien diferenciados y su expresión es menor en CCSC no diferenciados . Sin embargo, no está claro si CD109 se relaciona con invasión vascular, metástasis y pronóstico después de la cirugía. Por lo tanto, se necesitan más estudios para explorar la importancia clinicopatológica de CD109 en CCSC en un tamaño de muestra más grande.
CD109 es una glicoproteína ligada a GPI, que le permite ser liberada de la membrana . La forma soluble de CD109 también afecta la unión del TGF-β a sus receptores, y posteriormente modula la progresión del CCE . Litvinov et al. se encontró que CD109 liberado de la superficie celular en el medio extracelular, y la forma liberada de CD109 conserva su capacidad de inducir vías de señalización intracelular. Además, Sakakura et al. se informó que el suero CD109 fue liberado por el tumor xenoinjerto y aumenta proporcionalmente con el volumen del xenoinjerto tumoral. Por lo tanto, la detección de la concentración sérica de CD109 podría ayudar a vigilar los tumores que sobreexpresan CD109, incluidos los CCE.
Los exosomas comunican la lesión tumoral primaria y su nicho a través de su paquete que contiene proteínas seleccionadas u otras moléculas . El aislamiento y los análisis de exosomas circulantes relacionados con tumores pueden servir como biomarcadores para el diagnóstico de pacientes de cáncer. CD109 exógeno se ha identificado como un componente de exosoma secretado de células transfectadas de 293, por lo que es un objetivo prometedor para el diagnóstico de exosomas . Sin embargo, hasta la fecha no se han notificado CD109 en exosomas derivados de CCE u otras células tumorales. Los mecanismos subyacentes del envasado de CD109 en exosomas merecen una investigación más detallada.
Además del potencial como biomarcador, el CD109 podría ser un objetivo para abordajes terapéuticos. CD109 es una proteína de membrana, que puede ser dirigida directamente por anticuerpos o enzimas específicos. El CD109 también podría ser reconocido por vehículos específicos para la administración de drogas. Sin embargo, las funciones detalladas de CD109 en la patogénesis de los CCSC aún no están claras. Por ejemplo, es necesario definir la relación entre la expresión inferior de CD109 y los CCSC indiferenciados. La intervención con CD109 solo puede considerarse en la práctica clínica después de evaluar cuidadosamente los riesgos y beneficios.