
木葬による炭素隔離:基本的な評価
木葬による炭素隔離の可能性は、自然林には典型的に死んだ木が散らばっているという観察から生じている。 1). 更新世の氷期-間氷期の間に大北半球の氷床の下に大量の有機炭素が埋もれて保存されていたと仮定されている。 他の研究では、地方自治体の埋立地の有機物、特に木材は非常にゆっくりと分解することが示されています。 これらにより、木材の収穫と埋葬が炭素隔離のための実行可能な方法であり得ることが明らかになった。

自然な北アメリカの落葉性森林、ベルウッド、メリーランド州の林床に枯れ木。
世界的には、約60GtC y-1が土地植生によって一時的に隔離されています(純一次生産性またはNPP;図。 2). この炭素は、植生が死んで分解すると大気に連続的に戻されます(従属栄養呼吸、Rh)。 定常状態では、これらの炭素成分の死亡率はそれぞれの分解速度に等しく、正味の陸大気炭素フラックスがゼロに近い(NPP=Rh)ようにNPPまで加算される。 分解経路の一部を停止または減速させることができれば、現在の化石CO2排出量の8GtC y-1に匹敵する速度でCO2を隔離することが期待されます。 木質材料はリグニン-セルロース繊維構造のために分解に最も耐性があり、栄養ロックアップも最小限に抑えられているため(下)、この炭素プールに焦点を

2000-2006の人為的フラックスと1800-2006の累積プールを示す赤色のグローバル炭素循環の主要なプールとフラックスからのアップデートで、 地上生産性の約1/3(20GtC y-1)は木材生産であり、そのかなりの部分は木材埋葬による持続可能な炭素シンクの目標である。
この方法の可能性については、最初に2つの主要な質問に答える必要があります:枯れ木の生産率は何ですか? 残念なことに、林床の枯れ木に関する一般的な知識が不足しており、この炭素プールは炭素予算会計ではしばしば無視されています。 死亡率は基本的に成長率によって制限されるため、死んだ木材の生産率は60GtC y-1の世界総NPPを超えることはできません。 そして、重要な問題は、NPPが三つの主要な炭素プールにどのように分割されるかである:葉、木、根。 葉は成長し、毎年落葉性の森に落ちるが、常緑樹の森で数年続くことがあります。 小枝や小さな枝などの細かい木質材料は、しばしば壊れて落ちるかもしれませんが、木の幹や主要な枝は数十年から数世紀以上の寿命を持っています。 したがって、木質バイオマスは葉バイオマスよりもはるかに大きいにもかかわらず、その長い寿命は、他の方法よりもはるかに小さい生産率を示唆 根の生物量は大きい場合もあり、栄養素および水を捜すために根が絶えず育つと同時に死亡率はまた相当です。 「素朴な」最初の推測は、NPPがこれらの3つのプールに均等に分配され、20GtC y-1木材の成長率、したがって定常状態での20GtC y-1木材の死亡率につながる 微細な木質の破片はより迅速に分解され、取り扱いが困難になるため、幹や主要な枝などの粗い材料が埋葬に適しています。 木質材料の半分が粗いと仮定すると、約10GtC y-1死んだ木材が埋葬に利用可能であり、したがって10GtC y-1炭素シンクにつながる。 林床の枯れ木の平均滞留時間を10年と仮定すると、粗い木質の破片の形で約100GtC(10GtC y-1回10年)が林床に既に存在するであろう。 これらの死んだ木材は様々な崩壊段階にありますが、その半分を集めて埋めることができても、実質的に容易に入手可能な炭素シンクを提供します。
提案は、(1)林床に枯れ木を収集し、(2)生きている木を選択的に記録することです。 その後、木の幹は、林床に掘られたトレンチ(埋葬)または適切な埋立地に埋葬されるか、または雨から守られた地面の上に積まれた丸太のいずれかです(図。 3). 埋設された木質材料はかなり長い滞留時間を持ち、比較的速い分解プール(約10年)からはるかに遅い炭素プール(100-1000年以上)に効果的に炭素を移動させる。 (1)の場合、従属栄養呼吸の一部を減少させ、したがって即時有効な炭素シンクである。 (2)の場合、ツリーカットによって残された”ギャップ”におけるその後の再成長は、再成長の速度に依存する炭素シンクである。 実際には、(1)と(2)は、倒木が小さな木がケース(2)と非常によく似た方法で成長するためのギャップを残すので、おそらく大きな違いはありません。

森林の木の埋葬とストレージの概略図。
炭素隔離ポテンシャルの定量化
1木除去の持続可能なシンク(成長率によって制限される)
この潜在的な炭素シンクの大きさを定量化するために、グローバル動的植生と陸生炭素モデルVEGASを使用した。 このモデルは完全な地球炭素循環をシミュレートするが,ここでの目的に関連する炭素プールとフラックスのみを議論した。 シミュレーションは、このように推定値は、潜在的なレートになり、農地が含まれていませんでした。 モデルは、降水量、温度、日照、風速、蒸気圧の季節サイクルを持つ現代の観測された気候学によって駆動されました。 このシミュレーションは,樹木の成長が死亡率によってバランスされる定常状態で収束するまで実行した。モデル化されたグローバルNPPは57GtC y-1であり、そのうち19GtC y-1は枯葉に入り、17GtC y-1は枯木に入り、21GtC y-1は枯根構造に入る。 細かい木(小枝や小さな枝)はすぐに分解するので、取り扱いがより困難です(葉をきれいにするために高価ですなど)。)、およびより多くの埋葬スペースを占めるかもしれない、粗い木だけ埋葬のために適したように考慮されます。 林業の文献は一般に2つのクラスを分けるのに10cmの茎の直径を使用して良く、粗い木質の残骸の間で区別を、普通する。 残念なことに、細かい木と粗い木からの総木材死への相対的な寄与は、部分的には異なる寿命のために定量化することは困難である(小さな茎は、一般的に木全体よりも短い寿命を有する)。 これらのプールとフラックスがどのように定義されているのか、報告された数字が林業文献で何を表しているのかは不明な場合があります。 したがって、粗木材の死亡率が10GtC y-1になるように、死亡率のfine:coarse ratioを7:10にすることを幾分任意に指定します。
粗木材死亡率の空間分布を図に示す。 4. 最も高い割合は、アマゾンやコンゴ盆地などの熱帯雨林で見られ、続いて温帯林と北方林があります。 木材死亡率の空間分布が生産(NPP)の空間分布に似ているという事実は、死亡率が主に成長率に従うため、驚くべきことではありません。 3つのプール(葉:木:根=19:17:21)の間の全球平均分配比からの地域偏差は、植物機能タイプ(PFT)および気候依存炭素配分戦略の結果である。 このような偏差は、このモデルでは10-20%以下である。

kgC m-2y-1のモデルVEGASによって推定された世界の粗木材生産率。
さまざまな地理的地域の粗木材の炭素隔離ポテンシャルを表1に示します。 熱帯林は4.2GtC y-1炭素隔離ポテンシャルを有し、温帯林は3.7GtC y-1を有し、北方地域は2.1GtC y-1を有する。 このモデルは潜在的な植生のみを考慮しているため(農業は考慮していない)、温帯地域の潜在的な可能性は実質的に小さい可能性があります。
地域規模(表2)では、南米は2.3GtC y-1の炭素隔離ポテンシャルを持ち、アマゾンの熱帯雨林からの大きな貢献をしています。 アフリカは1.9GtC y-1に続いている。 ロシアは広大な北方の森林のために1.2GtC y-1の可能性を秘めています。 コンターミナスのUSは0の可能性があります。8GtC y-1は、東海岸と南に沿って広範な広葉樹と混合林、および山岳地帯の西に沿っています。 カナダは、その混合林と北方林から0.7GtC y-1の可能性を持っています。 中国の0.9GtC y-1の可能性のうち、国の森林の多くが長い間耕作地に変換されているため、おそらくほんの一部しか実現できません。 しかし、成功した森林再生プログラムは、この割合のサイズを高めることができます。
モデルによって推定された粗木材死亡率は、植物の機能タイプと気候に依存する炭素配分戦略の結果であり、現在のグローバル植生モデル この炭素プールとその回転率に関する観測は、一般的に欠けている。 それにもかかわらず、いくつかの研究では、この炭素プールの重要性が強調されています。 観察され、推定された平均樹死亡率を使用して、およびグローバルバイオマス分布を使用して点データを外挿すると、Harmon et al. 粗大な木質デブリの生産率は2-11GtC y-1と推定され、不確実性の範囲は木の寿命の推定値から来ています。 に基づいて、Matthewsは6GtC y-1を粗い木質の破片の生産率として推定しました。 比較を表3に示します。 したがって、VEGASモデルの結果は、の範囲内であるが、高い側にあります。 その理由の一つは、VEGASの平衡シミュレーションは、モデル化された森林が定常状態に達したこと、すなわち成熟した森林であることを意味し、使用されるデー 若い森林は古い成長の森林よりも死亡率が低い傾向があるため、成熟に向かって死亡率が増加するにつれて、これらの若い森林は将来的に高い可能性 両方の方法で多くの未知数が与えられた場合、2の不確実性の係数を10GtC y-1モデル推定値、すなわち5-15GtC y-1の範囲に割り当てます。
2既存の粗い木質の破片からの一度の可能性
過去の木の死の遺産として、腐敗の様々な段階で世界の森林にかなりの量の死んだ木が蓄積 5). 私はこの死んだ木のプールをシミュレートするためにモデルラスベガスを使用し、130GtCであることをグローバル粗い木質破片を推定しました,の75GtCの推定値 これらの数値は、この炭素プールに比較的ほとんど注意が払われていないため、大きく見えるかもしれませんが、世界の森林植生バイオマス(主に木材)に 崩壊の後期段階の木材は埋葬には適していないため(森林在庫調査に含まれる可能性も低い)、このプールの半分が埋葬に適していても、それはまだ65GtC 空間パターン(Fig. 5)は、主に低温での分解速度が遅いため、温帯および北方地域で高い値を持つ生産速度とは多少異なる分布を示しています。

kgc m-2における粗い木質デブリの世界分布。この大規模な既存の炭素プールの含意は、木材埋葬の初期段階では、上記の推定10GtC y-1の持続可能な速度以上が利用可能になるということです。
3生きた木の選択的切断による持続可能な率の向上
10GtC y-1枯木生産率は、積極的な森林管理によっても強化することができます。 木が死ぬのを待つのではなく、選択的な切断などの技術を介して比較的成熟した木を収穫することもできます。 一見すると、これは生きている木がCO2を取るので、炭素源のようです。 しかし、適切に木を選定すれば、若い森の方が生産性が高くなり、開発段階では生産性が呼吸や分解損失を大きく上回るため、全体的なシンクにつながる可能性がある。 生産性の低い木は光や他の資源のためにうまく競争しないので、それらの除去は若い木を隙間でより活発に成長させ、純炭素シンクを形成する。 偶数年の森林では、自己間伐は、若い木の大部分が他の木に道を譲るために死ぬ二次連続の主要なステップです。 この場合、多くの若い木は、死後に選択的に切断または収集することができます。
実装戦略
木材埋葬スキームの実装には、三つの主要なステップが含まれます:
- (1)
まだ場所にない場合は森林へのアクセスを可能にする;
- (2)
サイトの選択、埋葬のためのトレンチ掘削または地上貯蔵のためのシェルターを構築する;
- (3)
選択的な木の切断または死んだ木の収集に続いてトリミング、短縮、埋葬または貯蔵を適切なリターン間隔で繰り返す。私は、機械へのアクセスを可能にする道路や道のネットワーク、およびより均一ではない間隔で分散されるトレンチを想像しています。 例えば、1km×1kmの面積(100ヘクタール)は、典型的な粗木材生産率0.1kgC m-2y-1のために年間約100トンの炭素を蓄積するであろう(図。 4). 5年のリターン間隔で、各トレンチは500トンの炭素(約1000トンの乾燥木材塊)を埋めるだろう。 立方メートルあたり0.5トンの乾物を仮定し、丸太の間のスペースを無視すると、必要な容積は2000m3である。 杭が5メートルの土壌の下に埋設されている場合、トレンチは10m×10m×25mの寸法を有することができる(図。 6). 表面積は100m2であり、わずか0である。従って木製のコレクション区域の01%は、妨害小さいです。 土は丸太の間のそして上のスペースを満たし、解決するために許可される。 植生は、埋葬地に自然に戻って成長させることができます。 選択的な場所は埋められた木の腐食のために監視することができる。 図3と図6は、これらの手順を示しています。
図6 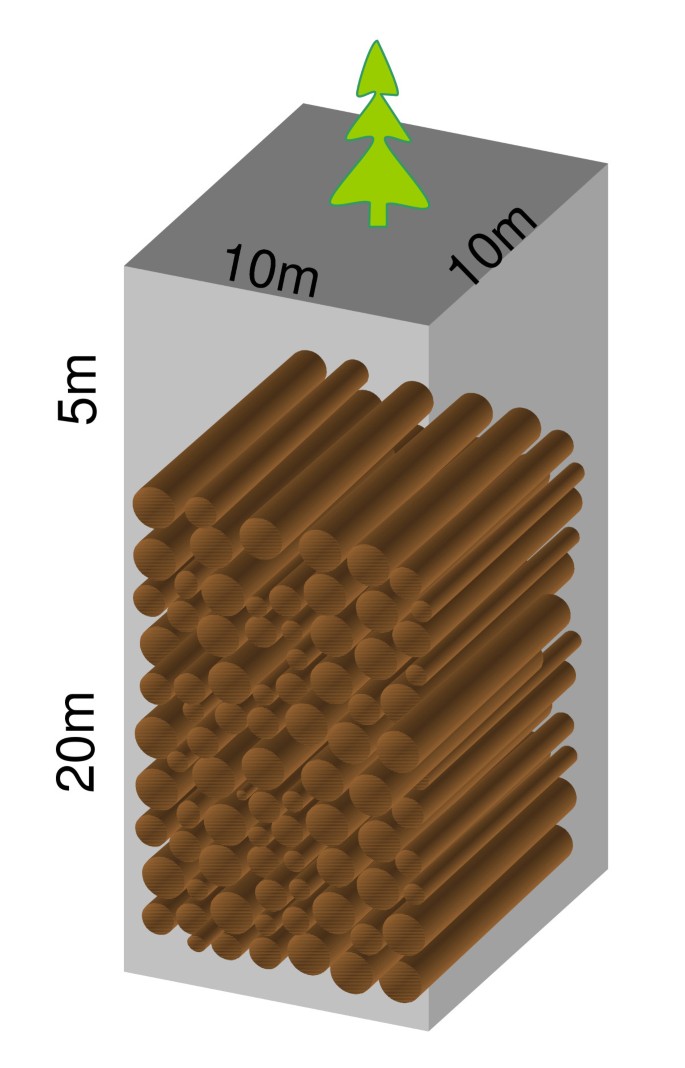
500tCを埋めることができるトレンチの例では、1km×1kmの典型的な中緯度森林面積からの粗木材炭素の量は5年間で500tCを埋めることができる。
実際のトレンチのサイズと分布は、トレンチの掘削、枯れ木の輸送、森林への外乱の最小化、分解を最も効果的に防止する場所の選択など、いくつかの要因のバランスをとる必要があります。 現地の埋葬は交通機関の費用を最小にするために可能な限り好まれます。 土が十分な深さの堀を掘るには余りにも浅いところで交通機関は必要かもしれません。 地形に伴う土壌水分量の変動など、小さな領域でも土壌状態が大きく変化する可能性があるため、サイトの選択に注意する必要があります。
死んだ木材の蓄積と腐敗率に応じて、このプロセスは数年(1-10)ごとに繰り返すことができますが、埋葬地は毎回異なります。 リターン間隔を選択するための主な基準は、各操作のコストと枯れ木が離れて腐敗させないようにする必要性との間のバランスになります。 自然の木の死がほとんどないように選択的切断が主な操作モードである場合(木は死ぬ前に切断される)、支配的な要因は除去するのに適した木の密度 プランテーションの場合は、低コストのために小さなセクション(グループカット)をクリアし、木が二次的な連続として成長することを可能にするのが良い戦略であるかもしれない。
地上の避難所の貯蔵と比較されて、堀の埋葬は分解の過程に普通既にあるので倒れた木のためのよりよい選択である、従って製材木としてより少なく有用である。 一方で、避難所の貯蔵は容易な使用のための製材木を将来の要求の増加べきである維持する。
木を収集または選択的に切断するために必要な技術は、ローテクであり、何千年もの周りされています。 現代の大規模な伐採のほとんどは、ヨーロッパや北米などの多くの場所で機械によって行われています。 アクセスのための道路システムは、米国の”森林高速道路”システムなど、これらの地域の多くですでに導入されています。 世界の森林の半分はすでに10km以内にあり、四分の三は主要な交通インフラから40km以内にあります。 大きな技術的ハードルがないので、このようなスキームは、これらの地域のかなりの部分でほぼすぐに実施することができます。 例えば、北米の林業の一般的な慣行は、私有または公有地で木を伐採するために、さまざまな操作スケールを持つ民間の伐採会社を雇うことであり、異な 現在激しく管理された森林に埋葬のためにすぐに利用できるほとんど死んだ木がないが、長期潜在性はまだ保持する。
このような分散システムは、炭素取引などのスキームを通じて経済的インセンティブが提供されている限り、監視を除いて、ほとんど政府の介入 北アメリカでは、森林に覆われた土地の多くは私有されています。 炭素隔離の可能性は、多くの地域の伐採業界と多くの土地所有者と経済にプラスの影響を与えるでしょう。 炭素吸収源の会計と監視は、伐採企業が収穫の各ラウンドのために戻ってくるときに認定されたエンジニアによって行うことができます。 これは、将来の衛星CO2観測によって支援された大気CO2測定を用いた渦相関フラックス測定、ソース/シンク反転などの大規模な監視システムによっ カナダとユーラシアの広大な北方林は、現在、部分的にアクセス可能であり、ほとんど管理されていないが、道路などのインフラは、関連する国では比較的迅速に建設することができる。
推定された10GtC y-1炭素隔離ポテンシャルの大部分を利用する場合、世界のほぼすべての森林を管理する必要があります。 次に、主な質問は、遠隔地の森林地域へのアクセス可能性です。 第一に、非常に急な山岳地帯や沼地の湿地へのアクセスが困難になります。 第二に、深い熱帯林への道はほとんどありません。 さらに、熱帯雨林の中心部に道路のネットワークを構築する提案は、生物多様性の損失などの大きな環境問題を引き起こすでしょう。 一方、経済的インセンティブは、このような道路の拡大を刺激していきます。 この場合であっても、違法な森林破壊のための法執行機関の問題、およびより広範なガバナンスの問題は、これらの地域の国がノーリターンに達する前に、最初に保証される必要があります。 近い将来、有益な練習は、継続的な森林破壊を伴う地域の木を燃やすのではなく、埋めることです。
熱帯雨林のコアをそのまま放置する場合、総炭素隔離ポテンシャルの約20%(熱帯雨林の半分)を占めています; 表1)、残りの熱帯、温帯および北方地域における隔離は、依然として8GtC y-1のシンクを提供する。 森林が通常よりよく保存されている急な地形にアクセスすることの難しさは、この数をさらに減らすでしょう。 実際には、道路建設のコストと環境への懸念を与えることは、単位面積当たりの炭素隔離率が低い大きな割合を乱すよりも、選択的な切断や細かい木質デブリの一部を埋めるなどの方法により、利用可能な森林のより小さな割合をより効率的に管理することが望ましい。
コスト
気候変動問題の規模は、代替エネルギー源、炭素隔離技術、または地理工学的アプローチであるかどうかにかかわらず、緩和戦略は、大規模に運用された場合に費用対効果がなければならないことを指示している。 米国の伐採業界からのデータによると、1トンの木材を収穫するための典型的なコストは約$20です。 木材は埋められる粗い木質材料の一部に過ぎないので、木材だけでは約50%以上であると仮定します(幹に比べてかなりの量の小さな枝があります)。 他の方向では、木材には水が含まれており、植物の乾燥質量が約50%の炭素であることを考えると、隔離された炭素のトン当たり40ドルになる可能性が これは溝の掘ること、道路工事および維持の費用が含まれていればより高いです。 したがって、隔離された1tC(トンまたは106グラムの炭素)のコストは$50になり、不確実性の範囲は2 25–tC100/tCになります。
これを発電所のCO2捕獲と地質学的貯蔵(CCS;表4)と比較することは、激しい研究の下で行われている戦略であることを示しています。 木材埋葬のためのcost50/tC($14/tco2)コストは、発電所CCSのための2 20-270/tco2よりも低いです。 発電所CCSの大きい範囲は安く利用できる古い鉱山がなくなると同時に増加する費用が原因である。 木製の埋葬の場合には、実用的な収蔵可能量の限定がありません。 産業用CCSの主なコストは、インフルエンザガスからのCO2の捕捉であり、木材埋葬は光合成の自然なプロセスによって行われるため、ゼロに近いコス
このコストを先駆的な欧州連合排出量取引システム(EUETS)の炭素キャップ-アンド-トレード市場価格と比較することも興味深い。 EUETSの価格は€1-33/tco2の間で変動しています2005-2007の間に。 これと比較して、自発的なシカゴ気候取引所(CCX)の価格は約$3-4/TCO2でした。 木材埋葬コストは現在の市場価格よりも幾分高いが、将来の気候緩和政策は炭素価格の上昇をもたらすと予想される。 地球規模で実施される場合、技術や人件費など、場所によって多くの要因が異なります。 最も安いのは、道路や機械が整備されている激しい管理下にある森林です。 この方法で利用される森林の総面積が増加するにつれて、価格が上昇する可能性があります。 機械の操作は、いくつかの化石燃料を消費し、CO2を放出します。 これらの要因を評価する必要があります。
木の埋葬と発電所CCSの表4の比較。 市場では炭素単位としてtco2を使用しており、変換係数は分子量比CO2:C=44:12でtCに変換することができ、両方の単位が示されています。 操作の規模
推定された電位(5GtC y-1)の半分だけが今後数十年で実行されたとしても、2050年までには、このような世界的な操作の規模
各トレンチが500tC容量を持っている場合(図の例。 6)、その後、5GtC y-1隔離率のために必要なトレンチの数は、年間10万人、すなわち3秒ごとに一つのトレンチになります。 10人の乗組員(機械付き)が1週間でトレンチを掘り、100ヘクタールの面積にわたって木材を収集/切断して埋めるのにかかると仮定すると、200,000人の乗組員(2万人の労働者)と機械のセットが必要になる。 この見積もりは確かに単純化されており、密集したまたは急傾斜した自然林で実行する場合、作業はかなり労働集約的になる可能性があります。
このような操作の規模は一見想像するのは難しいかもしれませんが、CO2問題の巨大な規模は、効果的な緩和戦略も同等の規模でなければならな 現在の8GtC y-1化石燃料の炭素排出率は、毎秒250tCに相当します。 木材の炭素含有量は化石燃料とほぼ同じであるため、木材の埋葬が化石燃料の排出に対抗する場合(潜在的に可能性があるように)、木材を埋葬する速度(質量または体積のいずれかの点で)は、化石燃料を燃やす速度に匹敵する必要がある。 木の埋葬がポートフォリオの一部として使用される場合、操作はそれに応じて縮小することができます。
この操作の妥当性は、経済的な観点からより容易に理解することができる。 木材埋葬のためのtC50/tCコストは、5GtC y-1隔離率で年間$250億に相当します。 これは、世界の総国内総生産(GDP)のわずか0.5%であり、projected48兆2006年には、予測された5-20%のGDPに比べて気候変動による潜在的な経済的被害。 200万人の労働者の年間2500億ドルのコストは、半分が機械やその他のコストであると仮定すると、労働者あたり62,500ドルを意味します。 明らかに、労働および機械費用は異なった国で非常に異なる場合もあります。 事業によって提供される雇用機会やその他の経済へのプラスの影響は、多くの地域、特に発展途上国で魅力的になります。
