Système amélioré de culture continue à petite échelle
Dans cette étude, nous montrons d’autres progrès et possibilités d’application d’un système de culture continue à petite échelle, précédemment développé par Klein et al. , pour un degré accru de parallélisation et une manipulation améliorée ainsi que la surveillance des réacteurs individuels. Les principaux aspects des modifications apportées au système comprennent une augmentation de l’ensemble des cuves de culture parallèles de 8 à 24 réacteurs et une diminution du volume utile de 10 à 6,5 mL. Le présent système consiste en outre en des couvercles sur mesure logeant quatre orifices fixes utilisés pour l’aération, l’alimentation en milieu, l’élimination du bouillon ainsi que l’inoculation ou le prélèvement (Fig. 1). Outre les quatre ports, une sonde DO en forme de tige optique est insérée à travers le couvercle pour la surveillance DO sans perturber le processus de culture, et remplace ainsi le point de capteur fluorescent d’oxygène de la configuration précédente. Le bain d’eau, qui dans la configuration précédente maintenait une température de culture constante, a été remplacé par un bloc chauffant en aluminium sur mesure, qui est fusionné à un agitateur de microplaques. Comme la version précédente du système de bioréacteur à petite échelle a été validée à l’aide de la levure de fission Schizosaccharomyces pombe, nous présentons ici la configuration de culture améliorée pour les cultures de S. cerevisiae.
Les étapes de fonctionnement de base, ainsi que les ajustements des taux de dilution en sélectionnant le diamètre du tube et le débit de pompe appropriés de la pompe d’afflux de fluide, ont été effectués comme décrit précédemment. Ici, le poids de la teneur en liquide de chaque bioréacteur a été déterminé gravimétriquement à la fin de la culture, permettant le calcul précis du taux de dilution respectif avec un 5.écart de 1%. Le bouillon de culture et la phase gazeuse du réacteur ont tous deux été évacués par le même orifice du couvercle du réacteur à l’aide de la pompe d’efflux (Fig. 1). Des débits de pompage d’efflux de 7,5 mL min−1 ont été utilisés pour toutes les expériences de culture. Le débit de la pompe d’efflux était largement supérieur au débit de la pompe d’alimentation, générant une légère pression négative à l’intérieur de la cuve de culture. Cette différence de pression a entraîné un afflux d’air par l’orifice d’aération. Le coefficient moyen de transfert de masse d’oxygène kLa atteint était de 110 h-1, ce qui permettait des niveaux de DO bien supérieurs à 30% de saturation tout au long du processus de culture. Le pH n’a pas été suivi en ligne ni contrôlé pendant la culture, car le milieu a été a priori ajusté à un pH de 6,0, ce qui a abouti à un pH final de 5,5 dans le bouillon de culture. Le pH a été mesuré en ligne sur une base quotidienne à partir de la sortie des réacteurs et après la récolte. Le pH est resté constant dès que l’état d’équilibre a été atteint et l’effluent du réacteur a montré un écart mineur de 0,1 unité de pH (données non présentées).
En explorant la production de 3 HP dans des chimiostats à petite échelle à différents taux de dilution dans des conditions limites de C
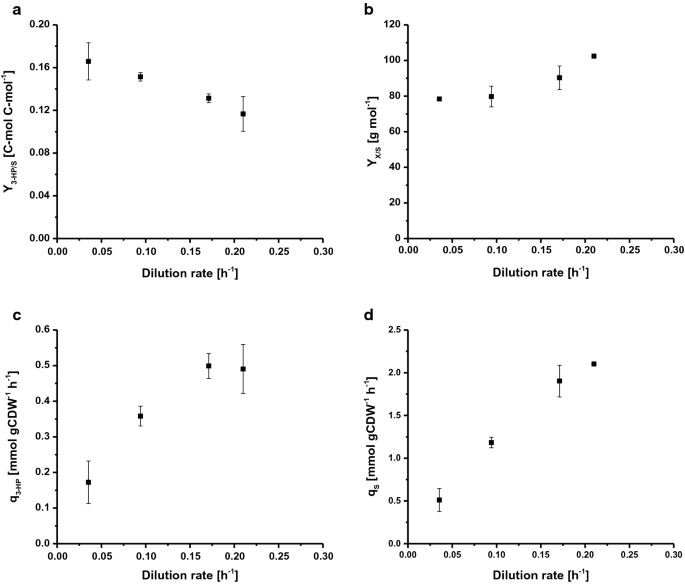
Afin de déterminer le taux de croissance spécifique maximal (µmax), S. cerevisiae ST938 a été cultivé dans des conditions de lot en appliquant une disponibilité excessive de nutriments. En utilisant le glucose comme source de carbone, le µmax était de 0,265 h-1, le rendement en biomasse était de 24,9 g mol−1 et le rendement en carbone de 3 HP sur le glucose était de 0,6% C-mol (tableau 1). Dans ce mode de culture, la majeure partie du carbone a été métabolisée en éthanol et en CO2, sous forme de flux glycolytiques élevés chez le type sauvage S. les cerevisiae sont fortement liées à la fermentation alcoolique. Les cultures de chimiostats sont contrôlées par l’apport d’un substrat limitant la croissance. À l’état d’équilibre, le S. cerevisiae sauvage ne produit pas de quantités significatives de métabolites de débordement en dessous d’un certain taux de croissance, marquant le taux de dilution critique (Dcrit (h−1)) en raison de l’absence d’accumulation de source de carbone. À ou au-dessus de ce taux de dilution critique, la source de carbone s’accumule dans le réacteur et le métabolisme de débordement est déclenché, entraînant la production de divers sous-produits, tels que l’éthanol, l’acétate et des quantités mineures d’acides organiques. En conséquence, des cultures continues ont été utilisées pour étudier la relation entre le taux de croissance et la formation de produit de 3 HP spécifique à la biomasse chez S. cerevisiae ST938. Les quatre taux de dilutions différents D(h-1) : 0,04, 0,09, 0,17 et 0,21 ont été choisis pour être inférieurs au µmax de la souche ST938 de S. cerevisiae déterminé en culture discontinue. Le rendement de la biomasse sur le glucose ainsi que le rendement de 3 HP, le taux de production spécifique de 3 HP et le taux d’absorption spécifique du substrat ont été étudiés (Fig. 2a-d, tableau 1). Tous les paramètres de culture pertinents, c’est-à-dire les rendements et les taux avec les écarts types respectifs, sont résumés dans le tableau 1.
Tableau 1 Paramètres de culture du S. cerevisiae ST938 cultivé en cultures continues dans des conditions C limitées
Fig. 2
Selected rates and yields for C-limited aerobic chemostat cultivations of S. cerevisiae ST938 a 3-HP yield (C-mol C-mol−1) on glucose, b biomass yield on glucose (g mol−1), c specific 3-HP production rate (mmol gCDW−1 h−1) and d specific glucose uptake rate (mmol gCDW−1 h−1) at different dilution rates for S. cerevisiae ST938. Cultivations were carried out in triplicates at 30 °C and pH 5.5 under C-limited conditions. Les erreurs correspondent à des écarts types dérivés de cultures triples
La culture de S. cerevisiae ST938 dans des conditions de limitation de C a montré un passage du métabolisme principalement fermentatif observé en mode batch à un métabolisme respiratoire chez les chimiostats, ce qui se traduit par des rendements de biomasse plus élevés ainsi que par une formation négligeable d’éthanol et de glycérol (tableau 1). De plus, des quantités mineures de glucose résiduel inférieures à 0.1 mM ont été détectés dans les échantillons prélevés à l’efflux des différents réacteurs, vérifiant que les cultures sont limitées en glucose. Les conditions limitées en carbone trouvées dans les cultures de chimiostats à l’état d’équilibre semblaient favoriser la formation de 3 HP, car les rendements de ce produit étaient 20 à 25 fois plus élevés que le rendement déterminé en culture discontinue (tableau 1). Cela est probablement dû à la conversion plus efficace du carbone en énergie sous forme d’ATP pendant la respiration par rapport à la fermentation dans des cultures discontinues. Fait intéressant, les cultures de S. cerevisiae ST938 dans des conditions de limitation de C a révélé qu’avec des taux de croissance spécifiques en baisse, une augmentation constante des rendements en carbone de 3 HP pouvait être observée, avec le rendement le plus élevé de 3 HP de 16,6% C-mol observé à un taux de dilution de 0,04 h-1 (Fig. 2 bis). Le rendement de 3 CV sur la biomasse est décuplé par rapport au rendement des cultures discontinues et a été multiplié par deux, passant de 0,19 à 0,43 g de gCDW−1, en abaissant le taux de dilution de 0,21 à 0,04 h−1 (tableau 1). La productivité spécifique maximale pour 3 HP d’environ 0,50 mmol gCDW-1 h-1 a été observée à des taux de dilution de 0,17 et 0.21 h-1, ce qui est environ quatre fois plus élevé que mesuré dans les cultures en lots (Fig. 2c, Tableau 1). Comme prévu, le taux d’absorption spécifique du glucose (qS) a augmenté avec des taux de dilution plus élevés de 0,51 à 2,1 mmol gCDW−1 h−1 (Fig. 2d). Ces valeurs sont jusqu’à 20 fois inférieures au taux d’absorption spécifique maximal de glucose de 10,7 mmol gCDW−1 h−1 observé à µmax dans des conditions de croissance limitées en glucose en mode batch (tableau 1). Des valeurs obtenues pour le taux d’absorption spécifique du substrat et le rendement de la biomasse à un taux de dilution de 0.09 h-1 (tableau 1) étaient en accord avec les données des cultures de chimiostat avec S. cerevisiae de type sauvage à un taux de dilution de 0,10 h−1, indiquant 1,1 et 1,25 mmol gCDW−1 h−1 ainsi que 0,49 g g−1, respectivement. Il a été constaté que le rendement en biomasse par substrat augmente avec l’augmentation des taux de dilution, pour le S. cerevisiae sauvage, cependant, ce paramètre est généralement constant en dessous du Dcrit tant que le métabolisme d’entretien constitue un puits de carbone considérable réduisant le rendement en biomasse à de faibles taux de dilution. En cas de S. souche cerevisiae ST938, cependant, il semblait que le carbone, qui n’était pas dirigé vers la formation de biomasse, contribuait dans une certaine mesure à la formation de 3 HP et de sous-produits.
Pour conclure, les différences observées dans les cultures continues à différents taux de dilution ainsi que par rapport à des configurations similaires avec le type sauvage de S. cerevisiae suggèrent que la voie biosynthétique intégrée pour produire 3-HP a un impact énorme sur la physiologie de la levure, qui est probablement amplifiée par les réponses adaptatives au stress. Enfin, C – culture de chimiostat limitée au taux de dilution le plus bas de 0.04 h-1 a donné le rendement en carbone le plus élevé de 3 CV.
Après avoir caractérisé S. cerevisiae ST938 dans des conditions de limitation de C, des cultures dans des conditions de limitation de l’azote et du phosphate ont été effectuées dans des conditions de limitation de l’azote et du phosphate, car elles se sont révélées auparavant favorables à la production de divers métabolites natifs et non natifs dans différents organismes. Ici, les cultures correspondantes sous limitation d’azote (N) et de phosphore (P) ont été étudiées dans le système de culture continue à petite échelle (Fig. 1) avec un taux de dilution fixé à 0,04 h-1, ce qui avait auparavant entraîné les rendements en carbone les plus élevés de 3 HP dans des conditions limitées en C (tableau 1). Les paramètres et les écarts types respectifs par rapport aux cultures limitées en N et en P de la souche de levure ST938 sont résumés dans le tableau 2.
Tableau 2 Paramètres de culture du S. cerevisiae ST938 cultivé en cultures continues à un taux de dilution de 0.04 h-1 dans des conditions limitées au carbone (C), à l’azote (N) et au phosphore (P)
Pour des conditions de culture limitées à N et P, aucun ammonium ou phosphate résiduel n’a été détecté dans les échantillons prélevés à l’efflux des différents réacteurs, vérifiant les cultures à limiter par rapport à la limitation correspondante du substrat (Tableau 2). Cependant, des quantités mineures de glucose résiduel de l’ordre de 0,63 et 0,19 mM ont été détectées sous limitation N et P (tableau 2). Au cours de la limitation C, les taux de glucose résiduel étaient bien inférieurs à 0,1 mM (tableau 2). Les cultures appliquant des limitations de N et de P ont révélé un rendement de 3 CV de 17,7 et 21,1% en mol de C, respectivement, affichant une valeur significativement plus élevée dans des conditions de limitation de P par rapport au rendement en carbone de 3 CV de 16,6 % en mol de C calculé pour des conditions de culture de limitation de C (tableaux 1, 2). De même, la productivité spécifique de la 3-HP a été significativement augmentée pour les cultures limitées en N et en P, révélant des valeurs de 0,22 et 0,23 mmol gCDW-1 h-1 par rapport à 0,17 mmol gCDW−1 h−1 atteintes dans les cultures limitées en C. De plus, les rendements de 3 CV sur la biomasse ont montré des valeurs comparables de 0,43 et 0.44 g de gCDW-1 pour les conditions limitant C et N, cependant, en cas de limitation P, un rendement accru de 0,50 g de gCDW-1 a été observé. En raison des faibles concentrations globales de sous-produits quantifiées, celles-ci sont négligeables (tableau 2). Les valeurs du taux d’absorption spécifique du substrat (qS) pour la limitation C et P étaient comparables et une valeur légèrement élevée de 0,61 mmol gCDW-1 h-1 a été obtenue dans des conditions N-limitées. Les rendements de biomasse par substrat ont révélé pour des conditions limitées à N et à P des valeurs similaires de 72,4 et 75,2 g de mol−1, respectivement. Par rapport au rendement en biomasse de 78.4 g mol−1 obtenus dans des conditions limitées en C, ces valeurs sont similaires. Notre étude soutient la justification selon laquelle des rendements de produits plus élevés par substrat ont été obtenus dans des conditions limitantes de N et de P. Par conséquent, il apparaît qu’un taux d’absorption spécifique du substrat plus élevé en cas de limitation de N combiné à la formation minimale de sous-produits a finalement favorisé la formation du produit. Dans des conditions de limitation de P, les moindres quantités de sous-produits se sont formées, ce qui pourrait favoriser la synthèse du produit. On peut en outre supposer que certains changements métaboliques peuvent entraîner une diminution de la quantité de carbone libérée sous forme de CO2.
En résumé, les cultures de chimiostat sous limitation de N et de P ont révélé des rendements accrus de 3 CV et des taux de production spécifiques par rapport aux conditions limitées en C, la limitation de P permettant les rendements les plus élevés.
Transférabilité des paramètres physiologiques mesurés dans des chimiostats à petite échelle vers des cultures en lots nourris dans des bioréacteurs de paillasse de 1 L dans des conditions limites de C et de P
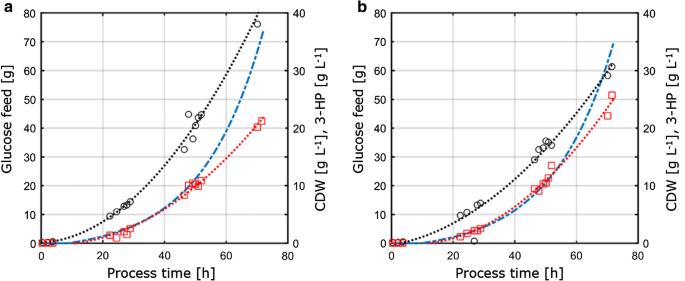
Comme cette étude visait davantage à évaluer la transférabilité et la comparabilité des paramètres physiologiques mesurés dans différentes conditions de culture, les paramètres de culture obtenus à partir des conditions limites de C et de P déterminées dans des cultures de chimiostats ont été transférés dans des réacteurs de paillasse agités de 1 L fonctionnant en mode lots nourris. Le concept de cette approche était donc de maintenir constantes les conditions et paramètres clés pour les cultures de chimiostat et de lots nourris afin d’assurer la comparabilité. Ces conditions comprenaient (i) le même rapport C: P que celui appliqué dans les cultures de chimiostats limitant le P, (ii) les conditions de processus identiques telles que le pH et la température, et (iii) les mêmes taux de croissance spécifiques que ceux appliqués dans les chimiostats, obtenus grâce à un profil d’alimentation exponentiel. Pour optimiser le titre de produit, les paramètres avec le rendement maximal en produit par substrat des expériences de chimiostat (D = 0.04 h-1) sous limitation C et P ont été choisis pour le transfert vers le système par lots alimentés. En raison de restrictions techniques dans la configuration, les cultures par lots nourris ont été effectuées à un taux de croissance de 0,05 h-1, ce qui est légèrement supérieur au taux de dilution défini de 0,04 h-1 dans les cultures de chimiostat. La culture en lots nourris a consisté en une phase initiale de lots pour générer de la biomasse suivie d’une phase d’alimentation exponentielle limitée en nutriments pour contrôler le taux de croissance. Seule la phase d’alimentation a été considérée comme pertinente pour l’évaluation de la transférabilité des paramètres obtenus à partir du système de culture continue, car uniquement pendant cette phase, la concentration du substrat contrôlait et limitait ainsi la croissance microbienne. Les profils de culture par lots nourris de S. cerevisiae ST938 appliquant des conditions de limitation C et P sont illustrés à la Fig. 3, et les paramètres de culture correspondants avec leur écart-type respectif sont résumés dans le tableau 3.
Fig. 3
Culture par lots aérobie dans des fermenteurs de banc de 1 L de S. cerevisiae ST938 en utilisant une rampe d’alimentation exponentielle à D = 0,05 h-1 avec le substrat limite a carbone, ou phosphore b. Les cercles noirs se réfèrent au titre de biomasse, les carrés rouges au titre de 3 HP et la ligne pointillée bleue à la quantité absolue de glucose introduite dans les réacteurs
Tableau 3 Paramètres de culture de S. cerevisiae ST938 cultivé en mode « fed-batch » dans des bioréacteurs de paillasse de 1 L dans des conditions limitantes du carbone (C) et du phosphore (P)
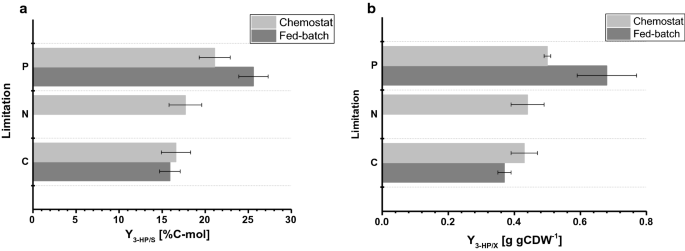
Le rendement en biomasse observé dans les cultures fed-batch était de 77,9 et 67,7 g de mol-1 pour la limitation C et P, respectivement. Le rendement en biomasse pour les cultures limitées en C était comparable entre le chémostat et l’exploitation par lots nourris, tandis que pour la limitation en P, un rendement en biomasse légèrement diminué a été observé pendant l’exploitation par lots nourris (tableaux 2 et 3), ce qui est dans les écarts-types observés pour les deux rendements en biomasse. Le rendement en carbone par substrat pour la 3-HP était de 15,9% pour la limitation en C, ce qui est presque identique par rapport à la valeur calculée dans le système de chimiostat (Fig. 4 bis). Les rendements de 3 HP déterminés dans cette étude sont en outre en accord étroit avec un rendement en carbone par substrat de 14% obtenu dans des études précédentes sur lots nourris à pH 5,0 avec une souche de S. cerevisiae conçue pour utiliser la voie de la β-alanine et de 13% avec un S. cerevisiae utilisant la voie dépendante de la malonyl-COA réductase. Des rendements plus élevés en carbone de 3 HP par substrat via la voie de la β-alanine ont été déterminés dans des études avec Escherichia coli indiquant une valeur de 42% dans les cultures nourries sur glucose.
Fig. 4
Comparaison des paramètres de culture sous différentes limitations déterminées dans des chimiostats à petite échelle et des réacteurs de paillasse de 1 L en mode par lots alimentés. a Rendement en carbone de 3 CV (% C-mol) et b rendement en biomasse de 3 CV (g gCDW-1). Les erreurs correspondent à des écarts types dérivés de cultures triples
Pour les conditions de limitation de P, cependant, notre étude a révélé une augmentation considérable du rendement en carbone de 3 CV de 25,6% (Fig. 4 bis). Les rendements de 3 HP observés sur la biomasse étaient respectivement de 0,38 et 0,65 g de gCDW-1 pour les conditions limitantes C et P. Pour la limitation C, cela a confirmé les valeurs observées dans la configuration du chimiostat (Fig. 4b). En raison de l’augmentation du rendement de 3 HP par substrat dans des conditions de limitation de P et de la légère diminution du rendement en biomasse par substrat en même temps, le rendement de 3 HP par biomasse a été significativement augmenté par rapport à l’expérience de chimiostat. Contrairement aux cultures effectuées en chémostats, aucune accumulation significative d’éthanol ou de glycérol n’a été détectée dans les cultures nourries par lots. Un bilan carbone fermé pour les conditions de limitation de C et de P indique qu’aucune quantité pertinente d’autres (sous-) produits n’a été formée (tableau 3). Cette différence dans le spectre des sous-produits pourrait expliquer l’augmentation du rendement de 3 CV par glucose.
En comparaison avec les cultures menées en chimiostats, les taux d’absorption spécifiques du glucose déterminés dans les cultures par lots nourris montrent des valeurs considérablement plus élevées de 0,75 et 0,77 mmol gCDW-1 h−1 pour les conditions limitantes C et P, respectivement. Comme mentionné ci-dessus, le point de consigne du facteur exponentiel du profil d’alimentation a été fixé à 0,05 h−1 et a varié jusqu’à 0,059 h−1, ce qui a entraîné un taux de croissance spécifique de 30 à 45% plus élevé dans les cultures nourries par lots par rapport aux expériences de chimiostat avec 0.04 h-1 (tableau 3), entraînant une augmentation des taux spécifiques d’absorption du glucose. Néanmoins, pour les fermentations limitées en C, le rendement en biomasse sur le glucose était comparable entre la culture par chémostat et la culture par lots nourris. De même, les taux de production spécifiques de 3 CV étaient élevés dans les cultures nourries par lots, avec 0,24 mmol gCDW-1 h−1 et 0,38 mmol gCDW−1 h−1, respectivement. Les rendements de 3 HP par substrat étaient comparables entre la culture par chémostat et la culture par lots nourris, ce qui suggère que les rendements sont un paramètre de culture robuste qui résiste aux perturbations induites par de plus petites fluctuations de la configuration de la culture. Aucun phosphate résiduel, ainsi que du glucose, n’a été détecté dans les échantillons prélevés dans les différents réacteurs pour des conditions de limitation de P, confirmant la limitation des cultures par rapport à la limitation correspondante. De même, pour la limitation de la C, aucun glucose résiduel n’a été détecté dans les échantillons.
Les améliorations présentées ici d’un système de chimiostat à petite échelle, précédemment développé par Klein et al. , comprenant entre autres une augmentation du nombre de réacteurs à 24 et une réduction du volume de culture à 6,5 mL. Ce degré accru de parallélisation fait du système un outil de criblage à haut débit approprié, car divers systèmes de chimiostat à petite échelle comparables fonctionnent avec des volumes de travail plus élevés et avec un nombre de réacteurs plus faible: Le système de bioréacteur à secousses parallèles continues (CosBios) utilise six vaisseaux de culture parallèles respectivement huit avec un volume de culture de 20 à 25 mL. Le système de bioréacteur à réservoir agité à usage unique modifié pour les cultures continues (bioréacteur, 2mag AG) fonctionne avec huit réacteurs parallèles à un volume de travail de 10 mL. Le système de mini-chimiostat (MC) développé par Bergenholm et al. comprend 16 réacteurs parallèles et nécessite un volume de travail de 40 mL. Notre système est donc bien applicable pour le criblage simple et rentable des performances microbiennes en mode continu. Il pourrait toutefois être pertinent d’étendre davantage le système en surveillant des paramètres supplémentaires, tels que les gaz d’échappement, pour permettre une analyse détaillée de la distribution du carbone, ce qui est particulièrement important pour les caractérisations physiologiques de la déformation.
Comme notre étude a suggéré que la limitation de N et de P était favorable à la formation de 3 HP chez S. cerevisiae, les expériences futures pourraient impliquer un examen plus approfondi des taux de croissance avec une limitation de N et de P appliquée afin de trouver la production optimale de 3 HP avec chaque limitation respective. Notre étude a également montré la transférabilité des paramètres physiologiques du chimiostat aux cultures nourries. Cela n’est possible que si aucun composé toxique ou inhibiteur ne s’accumule pendant la culture par lots nourris, car les paramètres physiologiques changeraient avec le temps et s’écarteraient des paramètres acquis dans les chimiostats. D’une manière générale, les expériences de chimiostat peuvent servir d’outil pour étudier l’influence d’un composé toxique ou inhibiteur potentiel en ajoutant la substance à l’aliment lui-même. Puisque tous les autres paramètres sont constants, l’influence du composé et sa concentration peuvent être directement évaluées et évaluées. Dans notre étude, aucune formation de sous-produits n’a été détectée dans les cultures en lots nourris et une réponse au stress vraisemblablement similaire a été induite en raison d’acides faibles, ce qui a permis aux conditions physiologiques des chimiostats à petite échelle de ressembler à celles des conditions en lots nourris.
En résumé, cette étude a montré que le concept de conditions de limitation de C et de P pour la production de 3 HP a été étudié dans un nouveau système de culture de chimiostat parallélisé et a pu être transféré avec succès à des bioréacteurs de paillasse de 1 L fonctionnant en mode par lots alimentés. Par conséquent, les paramètres physiologiques acquis dans les chimiostats peuvent être utilisés pour la conception et l’évaluation des performances des cultures nourries en utilisant des paramètres basés sur le rendement pour la configuration du processus.