
Introduction
Le renouvellement et la régénération des cellules ou des tissus sont les deux principales exigences du développement des organismes adultes. Les deux processus ont comme point de départ une population de cellules souches, normalement localisées dans un environnement spécifique appelé « niche », qui leur fournit les signaux requis pour maintenir les propriétés de la tige, ou pour se différencier des différents types de cellules requis (Figure 1). La prolifération et la différenciation des cellules souches doivent être coordonnées avec la mort des cellules à remplacer. En outre, des processus tels que la migration cellulaire, l’épigénétique et la communication cellulaire sont également nécessaires au renouvellement cellulaire approprié. Renouvellement rapideles tissus peuvent être reconnus par une activité mitotique plus élevée. Inversement, les tissus à renouvellement lent contiennent moins de mitose et peuvent ne pas être facilement reconnus dans les zones non renouvelées qui peuvent également présenter une mitose. Les décisions de devenir des cellules souches pendant la proliférationaffecter directement le renouvellement des tissus et l’homéostasie. Par conséquent, il est essentiel de comprendre les mécanismes de régulation qui soutiennent une division et une différenciation équilibrées des cellules. Les signaux extracellulaires (par exemple, le microenvironnement tissulaire, les ROS intracellulaires et les cytokines) ainsi que les facteurs intracellulaires (par exemple, les machines épigénétiques, les facteurs de transcription et la réponse aux dommages à l’ADN) sont responsables de la régulation de la division des cellules souches.
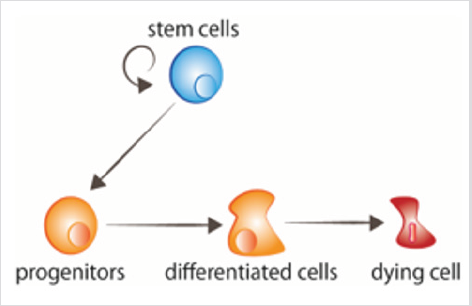
figure 1: Les cellules souches à renouvellement cellulaire prolifèrent, donnant naissance à des progéniteurs qui reçoivent ensuite les signaux de différenciation. Les cellules âgées reçoivent des signaux pour mourir.
Les cellules souches présentent trois options de division possibles: la division asymétrique, dans laquelle une cellule souche et une cellule fille engagée sont issues; l’engagement symétrique, qui regroupe deux cellules filles engagées; et la division symétrique, qui donne deux cellules filles qui conservent les propriétés des cellules souches Figure 2. Bien qu’on puisse prédire que la division asymétrique est le seul mécanisme qui permet le maintien d’une population stable de cellules souches, les données actuelles des expériences de recherche de lignées ont démontré que dans la plupart des tissus, l’équilibre entre la prolifération des cellules souches et la génération d’un ressort différentié est atteint au niveau de l’ensemble de la population de cellules souches.La perte de cellules souches due à la différenciation ou aux dommages cellulaires induitla division symétrique pour combler cette lacune. Après la division des cellules souches, les cellules qui suivent le processus de différenciation passent par différentes étapes définies par une combinaison de facteurs de transcription qui contrôlent l’activité du répertoire approprié de gènes et permettent leur engagement et leur différenciation terminale. Pour chaque lignage cellulaire, le produit final de la séquence de décisions est un type cellulaire différencié spécifique Figure 3a. Dans la plupart des circonstances, l’identité cellulaire – le produit de la différenciation normale – est stable dans les tissus et son maintien est crucial pour la fonction tissulaire normale. Une telle stabilité est obtenue grâce à une régulation épigénétique – par exemple, la déméthylation et l’acétylation des histones – qui se traduit par des modèles héritables d’expression génique spécifique aux tissus.
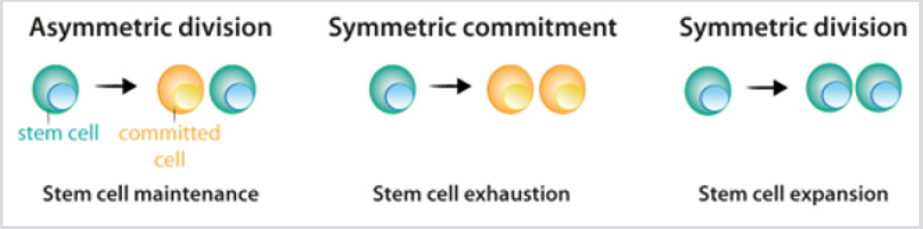
figure 2: Schéma de division des cellules souches.
a)Au cours de la division asymétrique, les cellules souches donnent naissance à une cellule souche, qui maintient la population de cellules souches, et une cellule qui devient engagée dans la différenciation.
b)Au cours de l’engagement symétrique, la division des cellules souches donne naissance à deux cellules filles qui se sont engagées àdifférenciation.
c)Lors de la division symétrique, la division des cellules souches donne naissance à deux cellules souches. Comme expliqué dans le texte, les données expérimentales actuelles indiquent que les trois modes de division peuvent se produire tout en maintenant la population de cellules souches.
Cependant, une perte d’identité de cellule peut se produire. En effet, les cellules du disque imaginal de Drosophile sont capables de transdéterminer et d’acquérir un nouveau destin d’adulte après la transplantation. Dans cette situation, les signaux extracellulaires semblent reprogrammer certains précurseurs ou cellules différenciées pour acquérir les caractéristiques d’un état plus stem ou d’un nouvel état différencié. Il existe deux mécanismes par lesquels une cellule peut changer son identité: la dédifférenciation et la transférenciation. La dédifférenciation désigne le processus par lequel une cellule différenciée ou engagée acquiert les caractéristiques d’une cellule moins mature Figure 3b. L’exemple le plus spectaculaire de la dédifférenciation est la conversion in vitro de cellules différenciées en phase terminale en cellules pluripotentes (cellules pluripotentes induites, IPSC), par surexpression d’un nombre limité de facteurs de transcription. La transdifférenciation, en revanche, se produit lorsqu’une cellule différenciée modifie son programme transcriptionnel et se convertit en un autre type de cellule différenciée. Le processus peut se produire par une étape intermédiaire de dédifférenciation vers un stade mature aless avant la conversion en la nouvelle cellule différenciée, ou directement, sans l’étape intermédiaire Figure 3c-d. La conversion directe de fibroblastes en myoblastes par l’expression ectopique de MyoD est un exemple du deuxième processus.
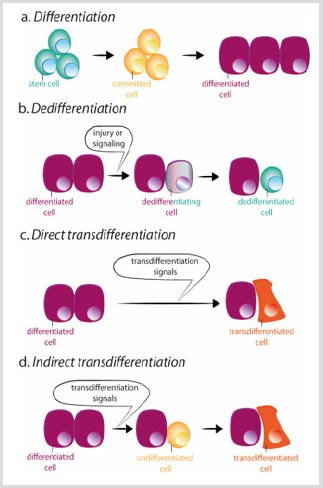
figure 3: Schémas de différenciation, de dédifférenciation et de différenciation trans.
a)Au cours de la différenciation normale, les cellules souches donnent naissance à des cellules engagées qui se différencient à leur tour en différents types de cellules.
b)La dédifférenciation consiste en l’acquisition des propriétés des cellules souches par une cellule différenciée. La différenciation trans peut se produirede manière directe ou indirecte.
c)Au cours de la différenciation trans directe, une cellule différenciée acquiert le programme transcriptionnel d’un autre type de cellule, généralement étroitement lié, comme par exemple exocrine aux cellules pancréatiques endocriniennes, devenant une cellule différenciée différente.
d)Lors de la différenciation trans indirecte, une cellule différenciée se dédifférencie avant d’adopter le nouveau programme transcriptionnel de l’autre type de cellule.
La dédifférenciation et la transdifférenciation se produisent également de manière naturelle en réponse à une blessure ou à une perte tissulaire.La dédifférenciation, par exemple, se produit naturellement lors de la limbergénération chez les amphibiens urodèles. Après l’amputation d’un membre, les cellules adjacentes à la plaie se dédifférencient, formant un blastème composé de cellules indifférenciées qui prolifèrent et finissent par se redifférencier dans le même type cellulaire pour créer tous les composants du membre perdu. La transdifférenciation naturelle se produit indirectement: d’abord, la cellule se dédifférencie; puis le programme de développement naturel est activé, permettant à la cellule de se différencier en une nouvelle lignée. Tsonis et ses collaborateurs ont décrit un naturelmécanisme de transdifférenciation dans un triton. Ils ont constaté que lorsqueles lentilles sont enlevées, les cellules épithéliales pigmentées de l’iris dorsaldifférencient et régénèrent le tissu manquant. Pour y parvenir, les cellules épithéliales pigmentées doivent d’abord se dédifférencier et se proliférer pour créer de nouvelles cellules cristallines, puis se différencier en cellules matures de la lentille. Dans les deux cas – la dédifférenciation et la redifférenciation dans le même type de cellule ou la transdifférenciation à un nouveau type de cellule – un réseau complexe de voies de signalisation peut contrôler le programme transcriptionnel acquis par chaque cellule dans le temps parfait.
le shu, le contrôle spatio-temporel de l’expression génique est continuellement requis pendant l’homéostasie animale et pendant le processus génératif. Cependant, pendant la régénération, les cellules doivent s’adapter à la nouvelle situation, ce qui nécessite de prendre des décisions plus solides au niveau cellulaire, y compris souvent des processus de différenciation et de transdifférenciation qui, pendant l’homéostasie, sont rares.
- Clevers H, Loh KM, Nusse R (2014) Signalisation des cellules souches. Un programme intégral pour le renouvellement et la régénération des tissus Signalisation Wnt et contrôle des cellules souches. Science 346 (6205): 1-9.
- Leblond C P, Walker B (1956) Renewal of cell population 36 (2): 255-276.
- Roostaee A, Benoit YD, Boudjadi S, Beaulieu JF (2016) Épigénétique dans le Renouvellement des Cellules épithéliales inTestinales. J Cell Physiol 231 (11): 2361-2367.
- Ito K, Ito K (2016) Metabolism and the Control of Cell Fate Decisions andStem Cell Renewal. Annu Rev Cell Dev Biol 32:399-409.
- Simons BD, Clevers H (2011) Strategies for homeostatic stem cell selfrenewalin adult tissues. Cellule 145(6): 851-862.
- Yang Y, Akinci E, Dutton JR, Banga A, Slack JM W, et al. (2013) Étapesprogrammation spécifique des cellules hépatiques d’embryons de souris en un phénotype de type cellule bêta. Mech Dev 130 (11-12): 602-612.
- Merrell AJ, Stanger BZ (2016) Plasticité des cellules adultes in vivo: la dédifférenciation et la transdifférenciation sont de retour à la mode. Nat Rev MolCell Biol 17 (7): 413-425.
- Worley MI, Setiawan L, Hariharan IK (2012) Régénération et détermination dans les disques imaginaux de la Drosophile. Annu Rév Genet 46:289-310.
- Raff M (2003) Plasticité des cellules souches adultes: Fait ou Artefact? Annu Rev CellDev Biol 19:1-22.
- Takahashi K, Yamanaka S (2006) Induction de Cellules Souches Pluripotentes à partir de Cultures de Fibroblastes Embryonnaires et Adultes de Souris par des facteurs définis. Cellule 126(4): 663-676.
- Tapscotr SJ, Davis RL, Thayer MJ, Cheng PF, Weintraub H, et al. (1988) MyoDi : Une Phosphoprotéine Nucléaire Nécessitant une Région d’homologie Myc pour convertir les Fibroblastes en Myoblastes. Science 242 (4877): 405-411.
- Jopling C, Boue S, Izpisua Belmonte JC (2011) Dédifférenciation, transdifférenciation et reprogrammation: trois voies de régénération.Nat Rev Mol Cell Biol 12 (2): 79-89.
- Tsonis PA, Madhavan M, Tancous EE, Del Rio-Tsonis K (2004) Del Rio-Tsonis, K. Vue oculaire d’un triton sur la régénération du cristallin. Int J Dev Biol 48 (8-9): 975-980.