
Protéines d’adhésion cellulaire
L’adhésion cellule-cellule est un processus sélectif, de sorte que les cellules n’adhèrent qu’à d’autres cellules de types spécifiques. Cette sélectivité a été démontrée pour la première fois dans des études classiques sur le développement embryonnaire, qui ont montré que les cellules d’un tissu (par exemple, le foie) adhèrent spécifiquement aux cellules du même tissu plutôt qu’aux cellules d’un tissu différent (par exemple, le cerveau). Cette adhésion cellule-cellule sélective est médiée par des protéines transmembranaires appelées molécules d’adhésion cellulaire, qui peuvent être divisées en quatre grands groupes : les sélectines, les intégrines, la superfamille des immunoglobulines (Ig) (ainsi nommées car elles contiennent des domaines structuraux similaires aux immunoglobulines) et les cadhérines (tableau 12.3). L’adhésion cellulaire médiée par les sélectines, les intégrines et les cadhérines nécessite du Ca2+ ou du Mg2+, de sorte que de nombreuses interactions adhésives entre les cellules sont dépendantes du Ca2+ ou du Mg2+.
Tableau 12.3
Molécules d’adhésion cellulaire.
Les sélectines médient les interactions transitoires entre les leucocytes et les cellules endothéliales ou les plaquettes sanguines. Il existe trois membres de la famille des sélectines: la L-sélectine, qui est exprimée sur les leucocytes; La E-sélectine, qui est exprimée sur les cellules endothéliales; et la P-sélectine, qui est exprimée sur les plaquettes. Comme indiqué précédemment dans ce chapitre, les sélectines reconnaissent les glucides de surface cellulaire (voir Figure 12.14). L’un de leurs rôles critiques est d’initier les interactions entre les leucocytes et les cellules endothéliales lors de la migration des leucocytes de la circulation vers les sites d’inflammation tissulaire (Figure 12.62). Les sélectines médient l’adhésion initiale des leucocytes aux cellules endothéliales. Ceci est suivi par la formation d’adhérences plus stables, dans lesquelles les intégrines à la surface des leucocytes se lient à des molécules d’adhésion intercellulaire (ICAMs), qui sont des membres de la superfamille Ig exprimés à la surface des cellules endothéliales. Les leucocytes fermement attachés sont alors capables de pénétrer dans les parois des capillaires et d’entrer dans le tissu sous-jacent en migrant entre les cellules endothéliales.

Figure 12.62
Adhérence entre les leucocytes et les cellules endothéliales. Les leucocytes quittent la circulation au niveau des sites d’inflammation tissulaire en interagissant avec les cellules endothéliales des parois capillaires. La première étape de cette interaction est la liaison des sélectines leucocytaires (plus…)
La liaison des ICAM aux intégrines est un exemple d’interaction hétérophile, dans laquelle une molécule d’adhésion à la surface d’une cellule (par exemple, une ICAM) reconnaît une molécule différente à la surface d’une autre cellule (par exemple, une intégrine). D’autres membres de la superfamille Ig médient des interactions homophiles, dans lesquelles une molécule d’adhésion à la surface d’une cellule se lie à la même molécule à la surface d’une autre cellule. Une telle liaison homophile conduit à une adhésion sélective entre cellules du même type. Par exemple, les molécules d’adhésion des cellules nerveuses (N-CAMs) sont des membres de la superfamille Ig exprimée sur les cellules nerveuses, et la liaison homophile entre N-CAMs contribue à la formation d’associations sélectives entre les cellules nerveuses au cours du développement. Il y a plus de 100 membres de la superfamille Ig, qui médient une variété d’interactions cellule-cellule.
Le quatrième groupe de molécules d’adhésion cellulaire, les cadhérines, présente également des spécificités de liaison homophiles. Ils ne sont pas seulement impliqués dans l’adhésion sélective entre les cellules embryonnaires, mais sont également principalement responsables de la formation de jonctions stables entre les cellules dans les tissus. Par exemple, l’E-cadhérine est exprimée sur les cellules épithéliales, de sorte que les interactions homophiles entre les E-cadhérines conduisent à l’adhésion sélective des cellules épithéliales les unes aux autres. Il est à noter que la perte d’E-cadhérine peut entraîner le développement de cancers provenant de cellules épithéliales, illustrant l’importance des interactions cellule-cellule dans le contrôle du comportement cellulaire. Différents membres de la famille de la cadhérine, tels que la N-cadhérine (cadhérine neurale) et la P-cadhérine (cadhérine placentaire), médient l’adhésion sélective d’autres types de cellules.
Une vingtaine de cadhérines classiques différentes, telles que l’E-cadhérine, ont été identifiées. De plus, une sous-famille distincte de cadhérines, appelées protocadhérines, est exprimée dans le système nerveux central où elles semblent jouer un rôle dans l’adhésion entre les neurones au niveau des synapses. Curieusement, différents neurones semblent exprimer différentes protocadhérines, suggérant que les protocadhérines peuvent jouer un rôle dans l’établissement de connexions spécifiques entre les neurones. Environ 50 gènes de protocadhérine humaine ont été identifiés et se sont organisés en trois groupes de gènes. Chaque amas contient plusieurs exons codant pour les domaines protocadhérine extracellulaire et transmembranaire N-terminaux, mais un seul ensemble de trois exons codant pour le domaine cytoplasmique C-terminal (Figure 12.63). Les amas de gènes de la protocadhérine semblent donc être constitués d’une région variable, codant de multiples domaines extracellulaires et transmembranaires, liée à une région constante codant un seul domaine cytoplasmique. Cette organisation des gènes de la protocadhérine ressemble de manière frappante à celle des gènes des immunoglobulines et des récepteurs des lymphocytes T (voir Figures 5.42 et 5.43), dans lesquels de multiples exons de régions variables sont joints à un seul exon de régions constantes. Dans les gènes des récepteurs des immunoglobulines et des lymphocytes T, cela résulte de réarrangements de l’ADN qui génèrent une diversité dans le système immunitaire. Il reste à déterminer si les régions variables et constantes des protocadhérines sont jointes au niveau de l’ADN ou de l’ARN (par exemple, par épissage alternatif) et dans quelle mesure des réarrangements de gènes de la protocadhérine pourraient contribuer à l’établissement de connexions synaptiques spécifiques dans le cerveau.
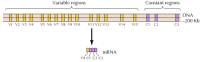
Figure 12.63
Organisation des groupes de gènes de la protocadhérine. Les gènes de la protocadhérine humaine sont organisés en trois groupes. Dans le cluster illustré, 15 régions variables différentes codant des domaines extracellulaires et transmembranaires sont liées à une seule région constante, (more…)
Contrairement aux jonctions stables cellule-matrice discutées dans la section précédente, les interactions cellule-cellule médiées par les sélectines, les intégrines et les membres de la superfamille Ig sont des adhérences transitoires dans lesquelles les cytosquelettes des cellules adjacentes ne sont pas liés les uns aux autres. Les jonctions d’adhésion stables impliquant les cytosquelettes des cellules adjacentes sont plutôt médiées par les cadhérines. Comme indiqué au chapitre 11, ces jonctions cellule-cellule sont de deux types : les jonctions adhérentes et les desmosomes, dans lesquels les cadhérines ou protéines apparentées (desmogléines et desmocollines) sont liées respectivement aux faisceaux d’actine et aux filaments intermédiaires (Figure 12.64). Le rôle des cadhérines dans la liaison des cytosquelettes des cellules adjacentes est donc analogue à celui des intégrines dans la formation de jonctions stables entre les cellules et la matrice extracellulaire.

Figure 12.64
Jonctions cellulaires stables médiées par les cadhérines. Les interactions homophiles entre les cadhérines médient deux types d’adhérences cellulaires stables. Dans les jonctions adhérentes, les cadhérines sont liées à des faisceaux de filaments d’actine via les caténines (voir Figure (plus…)