
Regulación y Control del Metabolismo en Bacterias(página 5)
(Este capítulo tiene 5 páginas)
© Kenneth Todar, PhD
Represión de Catabolitos
La inducción enzimática todavía se considera una forma de control negativo porque el efecto de la molécula reguladora (el represor activo) es reducir o reducir la tasa de transcripción. La presión cataboliteral es un tipo de control positivo de la transcripción, ya que una escritura reguladora afecta a un aumento (regulación ascendente) en la tasa de transcripción de un operón. El proceso fue descubierto en E. coli y se referíaoriginalmente como el efecto de la glucosa porque se descubrió que la glucosa reprimía la síntesis de ciertas enzimas inducibles, a pesar de que el inductor de la vía estaba presente en el medio ambiente. El descubrimiento se realizó durante el estudio de la regulación del operón lac en E. coli.Dado que la glucosa es degradada por enzimas constitutivas y la lactosa es degradada inicialmente por enzimas inducibles, ¿qué pasaría si la bacteria se cultivara en cantidades limitadas de glucosa y lactosa? Una gráfica de los evaluadores de crecimiento bacteriano resultó en una curva de crecimiento diáúxico que mostró dos fases distintas de crecimiento productivo (Figura 9). Durante la primera fase de crecimiento exponencial, la glucosa bacteriautiliza como fuente de energía hasta que toda la glucosa esté agotada.Luego, después de una fase de retraso secundaria, la lactosa se utiliza durante una segunda etapa de crecimiento exponencial.

Figura 9. La curva de crecimiento Diáúxica de E. coli cultivada en concentraciones limitantes de una mezcla de glucosa y lactosa
Durante el período de utilización de glucosa, la lactosa no se utiliza porque las células son incapaces de transportar y escindir el disacárido lactosa.La glucosa siempre se metaboliza primero en preferencia a otros azúcares. Solo después de que la glucosa se utiliza por completo, la lactosa se degrada. La lactoseoperona se reprime a pesar de la presencia de lactosa (el inductor). La razón ecológica es que la glucosa es una mejor fuente de energía que la lactosa, ya que su utilización requiere dos enzimas menos.
Solo después de agotar la glucosa se sintetizan las enzimas para la lactoseutilización. El retraso secundario durante el crecimiento diáxico representa el tiempo necesario para la inducción completa del operón lac y la síntesis de las enzimas necesarias para la utilización de la lactosa (permeasa de lactosa y beta-galactosidasa).Solo entonces se produce el crecimiento bacteriano a expensas de la lactosa. Dado que la disponibilidad de glucosa reprime las enzimas para la lactoseutilización, este tipo de represión se conoció como represión de catabolitos o el efecto de la glucosa.
Se sabe que la glucosa reprime un gran número de enzimas inducibles en diferentes bacterias. La glucosa reprime la inducción de las operaciones inductivas al inhibir la síntesis de AMP cíclico (cAMP), un nucleótido que se requiere para iniciar la transcripción de un gran número de sistemas de enzimas inducibles, incluido el operón lac.
El papel de un campamento cíclico es complicado. Se requiere cAMP para activar una proteína alostérica llamada CAP (proteína activadora de catabolitos)que se une al sitio de la CAP promotora y estimula la unión de la rnapolimerasa al promotor para iniciar la transcripción. Por lo tanto, para promover eficazmente la transcripción génica del operón lac, no solo debe estar presente la actosa para inactivar el represor lac, sino que el cAMP debe estar disponible para unirse al CAP que se une al ADN para facilitar la transcripción. Enla presencia de glucosa, la actividad de la adenilato ciclasa (AC) está bloqueada. Se requiere ACis para sintetizar cAMP a partir de ATP. Por lo tanto, si los niveles de cAMP son bajos, CAP está inactivo y no se produce la transcripción. Enla ausencia de glucosa, los niveles de cAMP son altos, CAP se activa por cAMP y se produce la transcripción (en presencia de lactosa).
Muchos promotores controlados positivamente, como el promotor lac, no son completamente funcionales en presencia de RNAp solo y requieren activación por CAP. El CAP está codificado por un gen Regulador separado, y está presente en niveles discretos. El CAP solo está activo en presencia del campamento. La unión de cAMP a CAP causa un cambio conformacional en la proteína permitiendo que se una al promotor cerca del sitio de unión de RNAp. CAP puede interactuar aparentemente con RNAp para aumentar la tasa de transcripción de operones en aproximadamente 50 veces.El control positivo del operón lac se ilustra en la Figura 10.
Figura 10. La presión catabolítica es un control positivo del operón lac. El efecto es un aumento en el ritmo de transcripción. En este caso, la proteína CAP se activa porcamppara unirse al operón lac y facilitar la unión de los polimeros de ARN al promotor para transcribir los genes para la utilización de lactosa.
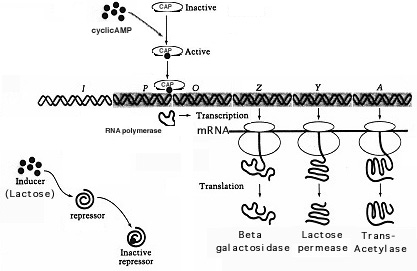
Como forma de represión de catabolitos, el efecto de la glucosa tiene una función útil en las bacterias: requiere que las células utilicen la mejor fuente de energía disponible. Para muchas bacterias, la glucosa es el sustrato más común y fácilmente utilizable para el crecimiento. Por lo tanto, inhibe indirectamente la síntesis de enzimas que metabolizan fuentes de energía más pobres.
FIN DEL CAPÍTULO
Página anterior
Volver a la página 1