
Zelladhäsionsproteine
Die Zell-Zell-Adhäsion ist ein selektiver Prozess, bei dem Zellen nur an andere Zellen bestimmter Typen haften. Diese Selektivität wurde erstmals in klassischen Studien zur Embryonalentwicklung nachgewiesen, die zeigten, dass Zellen aus einem Gewebe (z. B. Leber) spezifisch an Zellen desselben Gewebes und nicht an Zellen eines anderen Gewebes (z. B. Gehirn) haften. Eine solche selektive Zell-Zell-Adhäsion wird durch Transmembranproteine vermittelt, die als Zelladhäsionsmoleküle bezeichnet werden und in vier Hauptgruppen unterteilt werden können: die Selektine, die Integrine, die Immunglobulin (Ig) -Superfamilie (so genannt, weil sie immunglobulinähnliche Strukturdomänen enthalten) und die Cadherin (Tabelle 12.3). Die Zelladhäsion, die durch die Selektine, Integrine und Cadherin vermittelt wird, erfordert Ca2 + oder Mg2 +, so dass viele adhäsive Wechselwirkungen zwischen Zellen Ca2 + – oder Mg2 + -abhängig sind.
Tabelle 12.3
Zelladhäsionsmoleküle.
Die Selektine vermitteln transiente Wechselwirkungen zwischen Leukozyten und Endothelzellen oder Blutplättchen. Es gibt drei Mitglieder der Selectin-Familie: L-Selectin, das auf Leukozyten exprimiert wird; E-Selectin, das auf Endothelzellen exprimiert wird; und P-Selectin, das auf Thrombozyten exprimiert wird. Wie bereits in diesem Kapitel erläutert, erkennen die Selektine Kohlenhydrate an der Zelloberfläche (siehe Abbildung 12.14). Eine ihrer kritischen Aufgaben besteht darin, die Wechselwirkungen zwischen Leukozyten und Endothelzellen während der Migration von Leukozyten aus dem Kreislauf zu Stellen der Gewebeentzündung zu initiieren (Abbildung 12.62). Die Selektine vermitteln die anfängliche Adhäsion von Leukozyten an Endothelzellen. Es folgen stabilere Adhäsionen, bei denen Integrine auf der Oberfläche von Leukozyten an interzelluläre Adhäsionsmoleküle (iCams) binden, die Mitglieder der Ig-Superfamilie sind, die auf der Oberfläche von Endothelzellen exprimiert werden. Die fest gebundenen Leukozyten können dann die Wände der Kapillaren durchdringen und durch Migration zwischen den Endothelzellen in das darunter liegende Gewebe eindringen.

Abbildung 12.62
Adhäsion zwischen Leukozyten und Endothelzellen. Leukozyten verlassen den Kreislauf an Stellen der Gewebeentzündung, indem sie mit den Endothelzellen der Kapillarwände interagieren. Der erste Schritt in dieser Wechselwirkung ist die Bindung von Leukozytenselektinen (mehr…)
Die Bindung von iCams an Integrine ist ein Beispiel für eine heterophile Wechselwirkung, bei der ein Adhäsionsmolekül auf der Oberfläche einer Zelle (z. B. ein ICAM) ein anderes Molekül auf der Oberfläche einer anderen Zelle (z. B. ein Integrin) erkennt. Andere Mitglieder der Ig-Superfamilie vermitteln homophile Wechselwirkungen, bei denen ein Adhäsionsmolekül auf der Oberfläche einer Zelle an dasselbe Molekül auf der Oberfläche einer anderen Zelle bindet. Eine solche homophile Bindung führt zu einer selektiven Adhäsion zwischen Zellen des gleichen Typs. Beispielsweise sind Nervenzelladhäsionsmoleküle (N-CAMs) Mitglieder der Ig-Superfamilie, die auf Nervenzellen exprimiert werden, und die homophile Bindung zwischen N-CAMs trägt zur Bildung selektiver Assoziationen zwischen Nervenzellen während der Entwicklung bei. Es gibt mehr als 100 Mitglieder der Ig-Superfamilie, die eine Vielzahl von Zell-Zell-Interaktionen vermitteln.
Die vierte Gruppe von Zelladhäsionsmolekülen, die Cadherin, weisen ebenfalls homophile Bindungsspezifitäten auf. Sie sind nicht nur an der selektiven Adhäsion zwischen embryonalen Zellen beteiligt, sondern auch in erster Linie für die Bildung stabiler Verbindungen zwischen Zellen in Geweben verantwortlich. Beispielsweise wird E-Cadherin auf Epithelzellen exprimiert, so dass homophile Wechselwirkungen zwischen E-Cadherin zur selektiven Adhäsion von Epithelzellen aneinander führen. Es ist bemerkenswert, dass der Verlust von E-Cadherin zur Entwicklung von Krebserkrankungen führen kann, die aus Epithelzellen entstehen, was die Bedeutung von Zell-Zell-Interaktionen bei der Kontrolle des Zellverhaltens veranschaulicht. Verschiedene Mitglieder der Cadherin-Familie, wie N-Cadherin (neurales Cadherin) und P-Cadherin (plazentares Cadherin), vermitteln die selektive Adhäsion anderer Zelltypen.Etwa zwanzig verschiedene klassische Cadherin, wie E-Cadherin, wurden identifiziert. Darüber hinaus wird eine bestimmte Unterfamilie von Cadherin, genannt Protocadherin, im zentralen Nervensystem exprimiert, wo sie eine Rolle bei der Adhäsion zwischen Neuronen an Synapsen zu spielen scheinen. Interessanterweise scheinen verschiedene Neuronen unterschiedliche Protocadherin zu exprimieren, was darauf hindeutet, dass die Protocadherin eine Rolle bei der Herstellung spezifischer Verbindungen zwischen Neuronen spielen können. Etwa 50 menschliche Protocadherin-Gene wurden identifiziert und gezeigt, dass sie in drei Genclustern organisiert sind. Jeder Cluster enthält mehrere Exons, die für die N-terminalen extrazellulären und transmembranen Protocadherin-Domänen kodieren, aber nur einen einzigen Satz von drei Exons, die für die C-terminale zytoplasmatische Domäne kodieren (Abbildung 12.63). Die Protocadherin-Gencluster scheinen somit aus einer variablen Region zu bestehen, die für mehrere extrazelluläre und transmembrane Domänen kodiert und mit einer konstanten Region verbunden ist, die für eine einzelne zytoplasmatische Domäne kodiert. Diese Organisation von Protocadherin-Genen ähnelt auffallend der von Immunglobulin- und T-Zell-Rezeptorgenen (siehe Abbildungen 5.42 und 5.43), bei denen mehrere Exons variabler Region zu einem einzigen Exon konstanter Region verbunden sind. In den Immunglobulin- und T-Zell-Rezeptorgenen tritt dies als Folge von DNA-Umlagerungen auf, die Vielfalt im Immunsystem erzeugen. Es bleibt abzuwarten, ob die variablen und konstanten Regionen der Protocadherin auf DNA- oder RNA-Ebene verbunden sind (z. B. durch alternatives Spleißen) und inwieweit Umlagerungen von Protocadherin-Genen zum Aufbau spezifischer synaptischer Verbindungen im Gehirn beitragen können.
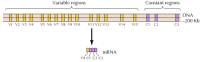
Abbildung 12.63
Organisation von Protocadherin-Genclustern. Die menschlichen Protocadherin-Gene sind in drei Clustern organisiert. In dem dargestellten Cluster sind 15 verschiedene variable Regionen, die extrazelluläre und transmembrane Domänen kodieren, mit einer einzigen konstanten Region verknüpft, (mehr…)
Im Gegensatz zu den im vorhergehenden Abschnitt diskutierten stabilen Zell-Matrix-Übergängen sind die durch die Selektine, Integrine und Mitglieder der Ig-Superfamilie vermittelten Zell-Zell-Wechselwirkungen vorübergehende Adhäsionen, bei denen die Zytoskelette benachbarter Zellen nicht miteinander verbunden sind. Stabile Adhäsionsübergänge, an denen das Zytoskelett benachbarter Zellen beteiligt ist, werden stattdessen durch die Cadherin vermittelt. Wie in Kapitel 11 erläutert, handelt es sich bei diesen Zell-Zell-Übergängen um zwei Arten: Adherens-Übergänge und Desmosomen, in denen Cadherin oder verwandte Proteine (Desmogleine und Desmocolline) an Aktinbündel bzw. Zwischenfilamente gebunden sind (Abbildung 12.64). Die Rolle der Cadherin bei der Verknüpfung des Zytoskeletts benachbarter Zellen ist somit analog zu der der Integrine bei der Bildung stabiler Verbindungen zwischen Zellen und der extrazellulären Matrix.

Abbildung 12.64
Stabile Zell-Zell-Verbindungen, vermittelt durch die Cadherin. Homophile Wechselwirkungen zwischen Cadherin vermitteln zwei Arten von stabilen Zell-Zell-Adhäsionen. In Adherens Junctions sind die Cadherin über die Catenine mit Bündeln von Aktinfilamenten verbunden (siehe Abbildung (mehr…)