
förbättrat småskaligt kontinuerligt odlingssystem
i denna studie visar vi ytterligare framsteg och tillämpningsmöjligheter för ett småskaligt kontinuerligt odlingssystem, som tidigare utvecklats av Klein et al. , för en ökad grad av parallellisering och förbättrad hantering samt övervakning av de enskilda reaktorerna. De viktigaste aspekterna av förändringarna i systemet innefattar en ökning av uppsättningen parallella odlingsfartyg från 8 till 24 reaktorer och en minskning av arbetsvolymen från 10 till 6,5 mL. Det nuvarande systemet består vidare av skräddarsydda lock som rymmer fyra fasta portar som används för luftning, mediaförsörjning, avlägsnande av buljong samt inokulering eller provtagning (Fig. 1). Förutom de fyra portarna sätts en optisk stavformad do-sond in genom locket för DO-övervakning utan att störa odlingsprocessen och på så sätt ersätta den syre-fluorescerande sensorfläcken i den tidigare inställningen . Vattenbadet, som i den tidigare uppställningen upprätthöll en konstant odlingstemperatur, ersattes av ett skräddarsytt aluminiumvärmeblock, som smälts till en mikroplattblandare. Eftersom den tidigare versionen av det småskaliga bioreaktorsystemet validerades med hjälp av Fissionjäst Schizosaccharomyces pombe , presenterar vi här den förbättrade odlingsuppsättningen för S. cerevisiae-odlingar.
grundläggande driftssteg, liksom justeringar av utspädningshastigheter genom att välja lämplig rördiameter och pumphastighet för mediatillströmningspumpen, utfördes som tidigare beskrivits . Här bestämdes vikten av vätskeinnehållet i varje bioreaktor gravimetriskt vid slutet av odlingen, vilket möjliggjorde den exakta beräkningen av respektive utspädningshastighet med en 5.1% avvikelse. Odlingsbuljong och reaktorgasfasen avlägsnades båda genom samma port på reaktorlocket med hjälp av utflödespumpen (Fig. 1). Utflödespumphastigheter på 7,5 mL min – 1 användes för alla odlingsexperiment. Utflödespumphastigheten var långt över matningspumphastigheten, vilket genererade ett litet negativt tryck inuti kulturkärlet. Denna tryckskillnad resulterade i inflödet av luft genom luftningsporten. Den genomsnittliga syremassöverföringskoefficienten kla uppnåddes var 110 h−1, vilket tillät DO-nivåer långt över 30% mättnad under hela odlingsprocessen. PH övervakades inte online eller kontrollerades under odlingen, eftersom mediet a priori justerades till ett pH på 6,0, vilket resulterade i ett slutligt pH på 5,5 i odlingsbuljongen. PH mättes i linje dagligen från utflödet av reaktorerna och efter skörd. PH förblev konstant så snart steady-state uppnåddes och reaktorutflödet visade en mindre avvikelse på 0,1 pH-enheter (data visas inte).
Utforska 3-HP-produktion i småskaliga kemostater med olika utspädningshastigheter under C-begränsande förhållanden
för att bestämma den maximala specifika tillväxttakten (jacobmax) odlades S. cerevisiae ST938 i batchförhållanden med användning av överskott av näringsämnen. Genom att använda glukos som kolkälla, den var 0,265 H−1, biomassautbytet var 24,9 g mol−1, och 3-hk kolutbytet på glukos var 0,6% C-mol (Tabell 1). I detta odlingsläge metaboliserades det mesta av kolet till etanol och CO2 , som höga glykolytiska flöden i vildtyp S. cerevisiae är starkt kopplade till alkoholjäsning . Kemostatodlingar styrs av tillförseln av ett tillväxtbegränsande substrat. Vid steady state producerar vildtyp S. cerevisiae inte signifikanta mängder överflödesmetaboliter under en viss tillväxthastighet, vilket markerar den kritiska utspädningshastigheten (Dcrit (h−1)) på grund av bristen på ackumulering av kolkälla. Vid eller över denna kritiska utspädningshastighet ackumuleras kolkälla i reaktorn och överflödesmetabolism utlöses, vilket resulterar i produktion av olika biprodukter, såsom etanol, acetat och mindre mängder organiska syror . Följaktligen användes kontinuerliga kulturer för att undersöka förhållandet mellan tillväxttakten och biomassa-specifik 3-HP-produktbildning i S. cerevisiae ST938. De fyra olika utspädningshastigheterna D (h-1): 0,04, 0,09, 0,17 och 0,21 valdes för att ligga under den enligt S. cerevisiae stamstam st938 bestämd vid satsodling. Biomassautbytet på glukos såväl som 3-hk-utbytet, specifik 3-hk produktionshastighet och den specifika substratupptagningshastigheten undersöktes (Fig. 2a-D, Tabell 1). Alla relevanta odlingsparametrar, dvs. utbyten och satser med respektive standardavvikelser, sammanfattas i Tabell 1.
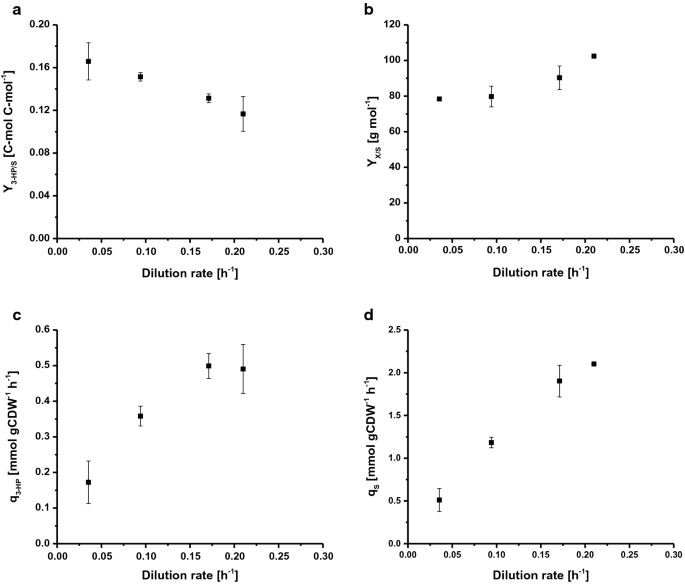
Selected rates and yields for C-limited aerobic chemostat cultivations of S. cerevisiae ST938 a 3-HP yield (C-mol C-mol−1) on glucose, b biomass yield on glucose (g mol−1), c specific 3-HP production rate (mmol gCDW−1 h−1) and d specific glucose uptake rate (mmol gCDW−1 h−1) at different dilution rates for S. cerevisiae ST938. Cultivations were carried out in triplicates at 30 °C and pH 5.5 under C-limited conditions. Fel motsvarar standardavvikelser som härrör från tredubbla odlingar
odlingen av S. cerevisiae ST938 under C-begränsande förhållanden visade en övergång från övervägande fermentativ metabolism observerad i batchläge till en respiratorisk metabolism i kemostater, vilket återspeglas i högre biomassaavkastning såväl som i försumbar etanol-och glycerolbildning (Tabell 1). Dessutom mindre mängder kvarvarande glukos under 0.1 mM detekterades i proverna som tagits från utflödet från de olika reaktorerna, vilket verifierade att kulturerna var glukosbegränsade. Kolbegränsade förhållanden som hittades i kemostatkulturer vid steady-state verkade främja bildandet av 3-hk, eftersom utbytet av denna produkt var 20 – till 25-faldigt högre än utbytet bestämt i batchkultur (Tabell 1). Detta beror troligen på den effektivare omvandlingen av kol till energi i form av ATP under andning jämfört med jäsning i batchkulturer. Intressant, odlingarna av S. cerevisiae ST938 under C-begränsande förhållanden avslöjade att med minskande specifika tillväxthastigheter kunde en konstant ökning av 3-hk kolutbyten observeras, med det högsta 3-hk-utbytet på 16,6% C−mol observerat vid en utspädningshastighet på 0,04 h-1 (Fig. 2a). 3-HP-utbytet på biomassa är tiofaldigt högre än utbytet i batchkulturer och ökade mer än dubbelt från 0,19 till 0,43 g gCDW−1 genom att sänka utspädningshastigheten från 0,21 till 0,04 h−1 (Tabell 1). Den maximala specifika produktiviteten för 3-HK på cirka 0,50 mmol gCDW−1 h−1 sågs vid utspädningshastigheter på 0,17 och 0.21 h-1, vilket är ungefär fyrfaldigt högre än mätt i satsodlingar (Fig. 2C, Tabell 1). Som förväntat ökade den specifika glukosupptagningshastigheten (qS) med högre utspädningshastigheter från 0, 51 till 2, 1 mmol gCDW−1 h−1 (Fig. 2d). Dessa värden är upp till 20 gånger lägre än den maximala specifika glukosupptagningshastigheten på 10, 7 mmol gCDW-1 h−1 observerad vid sackarimax under glukosbegränsade tillväxtförhållanden i batchläge (Tabell 1). Värden erhållna för den specifika substratupptagningshastigheten och utbytet av biomassa med en utspädningshastighet på 0.09 h – 1 (Tabell 1) överensstämde med data från kemostatodlingar med vildtyp S. cerevisiae vid en utspädningshastighet på 0,10 h-1, med angivande av 1,1 och 1,25 mmol gCDW−1 h−1 såväl som 0,49 G G−1 . Det har visat sig att biomassautbytet per substrat ökar med ökande utspädningshastigheter, för vildtyp S. cerevisiae är emellertid denna parameter i allmänhet konstant under Dcrit så länge underhållsmetabolism utgör en avsevärd kolsänka som minskar biomassautbytet vid låga utspädningshastigheter . I händelse av S. cerevisiae-stammen ST938 verkade emellertid att kol, som inte var riktat mot biomassbildning, i viss utsträckning bidrog till bildandet av både 3-HK och biprodukter.
För att avsluta, skillnader som upplevs i kontinuerliga kulturer vid olika utspädningshastigheter såväl som i jämförelse med liknande uppställningar med S. cerevisiae vildtyp tyder på att den integrerade biosyntetiska vägen för att producera 3-HP har en enorm inverkan på jästfysiologi, vilket förmodligen förstärks av adaptiva stressresponser. Slutligen C-begränsad kemostatodling vid den lägsta utspädningshastigheten på 0.04 h – 1 resulterade i det högsta 3-hk-kolutbytet.
undersökande kväve-och fosfatbegränsning för ökade 3-HP-utbyten i småskaliga kemostatkulturer
Efter karakterisering av S. cerevisiae ST938 under C-begränsande förhållanden utfördes odlingar under kväve – och fosfatbegränsande förhållanden, eftersom de tidigare har visat sig vara gynnsamma för produktion av olika infödda och icke – infödda metaboliter i olika organismer . Här undersöktes motsvarande odlingar under kväve (N) och fosfor (P) begränsning i det småskaliga kontinuerliga odlingssystemet (Fig. 1) med en bestämd utspädningshastighet på 0,04 h-1, vilket tidigare hade resulterat i de högsta kolutbytena på 3 hk under C-begränsade förhållanden (Tabell 1). Parametrarna och respektive standardavvikelser från n-och P-begränsade odlingar av jäststam ST938 sammanfattas i Tabell 2.
för odlingsförhållanden med N – och P-Begränsningar detekterades inget kvarvarande ammonium eller fosfat i de prover som tagits från utflödet från de olika reaktorerna, vilket verifierade de kulturer som ska begränsas med avseende på motsvarande substratbegränsning (Tabell 2). Emellertid detekterades mindre mängder kvarvarande glukos i intervallet 0,63 och 0,19 mM under n – och P-begränsning (Tabell 2). Under C-begränsning var de återstående glukosnivåerna långt under 0,1 mM (Tabell 2). Odlingar som tillämpar n-och P-Begränsningar avslöjade ett 3-hk-utbyte på 17,7 respektive 21,1% C-mol, vilket visar ett signifikant högre värde under P-begränsade förhållanden jämfört med 3-hk kolutbyte på 16,6% C-mol beräknat för C-begränsade odlingsförhållanden (tabeller 1, 2). På samma sätt ökades den specifika produktiviteten för 3-hk signifikant för n – och P-begränsade odlingar som avslöjade värden på 0,22 och 0,23 mmol gCDW−1 h−1 jämfört med 0,17 mmol gCDW−1 h−1 uppnådd i C-begränsade kulturer. Dessutom visade 3-HP-avkastningen på biomassa jämförbara värden på 0.43 och 0.44 g gCDW – 1 för C – och N-begränsande förhållanden observerades emellertid ett ökat utbyte av 0,50 g gCDW-1 vid p−begränsning. På grund av de totala låga koncentrationerna av biprodukter som kvantifierats är dessa försumbara (Tabell 2). Värdena för den specifika substratupptagningshastigheten (qS) för C – och P-begränsning var jämförbara och ett något förhöjt värde på 0,61 mmol gCDW−1 h−1 erhölls under n-begränsade betingelser. Biomassautbyten per substrat avslöjade för N – och P-begränsade förhållanden liknande värden på 72,4 respektive 75,2 g mol−1. Jämfört med biomassautbytet på 78.4 g mol – 1 uppnås under C-begränsade förhållanden, dessa värden är likartade. Vår studie stöder motiveringen att högre produktutbyten per substrat uppnåddes under N – och P-begränsande förhållanden. Följaktligen verkar det som om en högre specifik substratupptagningshastighet vid n-Begränsning kombinerad med minimal bildning av biprodukter i slutändan gynnade produktbildning. Under p-begränsande förhållanden bildades de minsta mängderna biprodukter, vilket skulle kunna gynna produktsyntes. Det kan vidare spekuleras i att vissa metaboliska förändringar eventuellt leder till att en sänkt mängd kol frigörs som CO2.
Sammanfattningsvis visade kemostatodlingar under n-och P-begränsning ökade 3-HP-utbyten och specifika produktionshastigheter jämfört med C-begränsade förhållanden, med P-begränsning som möjliggör de högsta produktutbytena.
överförbarhet av de fysiologiska parametrarna uppmätta i småskaliga kemostater till Fed-batchkulturer i 1-L bänkbioreaktorer under C – och P-begränsande förhållanden
eftersom denna studie vidare syftade till att bedöma överförbarhet och jämförbarhet av de fysiologiska parametrarna uppmätta under olika odlingsförhållanden, överfördes odlingsparametrarna erhållna från C – och P-begränsande förhållanden bestämda i kemostatkulturer till 1-L omrörda bänkreaktorer som körs i fed-batch-läge. Konceptet med detta tillvägagångssätt var därför att hålla viktiga förhållanden och parametrar konstanta för kemostat-och Fed-batchkulturer för att säkerställa jämförbarhet. Dessa villkor omfattade (i) samma C:P-förhållande som tillämpas i P-begränsande kemostatodlingar, (ii) identiska processförhållanden som pH och temperatur, och (iii) samma specifika tillväxthastigheter som tillämpas i kemostater, uppnås genom en exponentiell foderprofil. För att optimera produkttitern, parametrarna med maximal produktutbyte per substrat från kemostatexperimenten (D = 0.04 h-1) under C – och P-begränsning valdes för överföring till fed-batch-systemet. På grund av tekniska begränsningar i uppställningen utfördes matade satsodlingar med en tillväxttakt på 0,05 h-1, vilket är något högre jämfört med den inställda utspädningshastigheten på 0,04 h-1 i kemostatodlingar. Fed-batch-odlingen bestod av en initial satsfas för att generera biomassa följt av en exponentiell, näringsbegränsad utfodringsfas för att kontrollera tillväxthastigheten. Endast utfodringsfasen ansågs relevant för bedömningen av överförbarheten av parametrarna erhållna från det kontinuerliga odlingssystemet, eftersom substratkoncentrationen enbart under denna fas kontrollerade och därmed begränsade mikrobiell tillväxt. De fed-batch odling profiler av S. cerevisiae ST938 tillämpa C-och P-begränsande betingelser visas i Fig. 3, och motsvarande odlingsparametrar med respektive standardavvikelse sammanfattas i tabell 3.
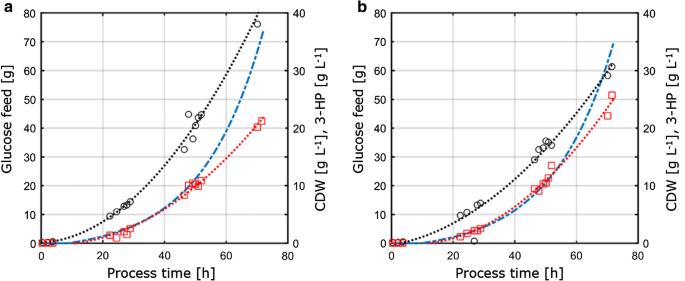
aerob fed-batch odling i 1-L bänk-top fermentorer av S. cerevisiae ST938 med användning av en exponentiell matningsramp vid D = 0,05 h−1 med det begränsande substratet a kol, eller B fosfor. Svarta cirklar hänvisar till biomassa titer, röda rutor till 3-hk titer och blå streckad linje till den absoluta mängden glukos som matas till reaktorerna
biomassautbytet som observerades i fed-batchkulturerna var 77,9 respektive 67,7 g mol−1 för C – respektive P-begränsning. Biomassautbytet för C-begränsade odlingar var jämförbart mellan kemostat och fed-batch-drift, medan för P-begränsning observerades en något minskad biomassautbyte under fed-batch-Drift (tabeller 2, 3), vilket ligger inom de standardavvikelser som observerats för båda biomassautbytena. Kolutbytet per substrat för 3-HK var 15,9% för C-begränsning, vilket är nästan identiskt jämfört med värdet beräknat i kemostatsystemet (Fig. 4a). De 3-HP-avkastningar som bestämts i denna studie är vidare i nära överensstämmelse med ett kolutbyte per substrat på 14% erhållet i tidigare C-begränsade fed-batchstudier vid pH 5.0 med en S. cerevisiae-stam konstruerad för att använda Baccarat-alanin-vägen och av 13% med en S. cerevisiae som använder den malonyl-CoA-reduktasberoende vägen . Högre 3-HP-kolutbyten per substrat via bronkialalanin-vägen bestämdes i studier med Escherichia coli som anger ett värde på 42% i fed-batch-odlingar på glukos .
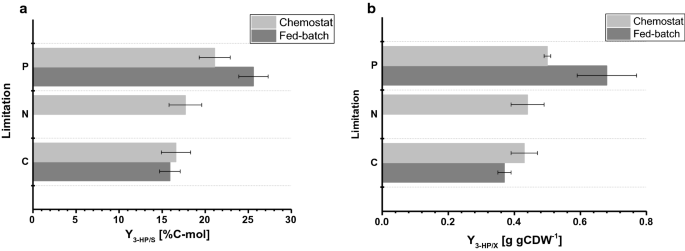
jämförelse av odlingsparametrar under olika begränsningar bestämda i småskaliga kemostater och 1-L bänkreaktorer i fed-batch-läge. en 3-hk kolutbyte (%C-mol) och B 3-hk utbyte på biomassa (g gCDW−1). Fel motsvarar standardavvikelser härledda från tredubbla odlingar
För p-begränsande förhållanden visade vår studie dock en avsevärd ökning av 3-hk kolutbyte på 25,6% (Fig. 4a). De observerade 3-hk-avkastningarna på biomassa var 0,38 och 0,65 g gCDW−1 för C – respektive P-begränsande förhållanden. För C-begränsning bekräftade detta de värden som observerades i kemostatuppsättningen (Fig. 4b). På grund av ökningen av utbytet av 3 hk per substrat under P-begränsande förhållanden och den lilla minskningen av biomassautbytet per substrat samtidigt ökade utbytet av 3 hk per biomassa signifikant jämfört med kemostat-experimentet. I motsats till de odlingar som utfördes i kemostater detekterades ingen signifikant ackumulering av etanol eller glycerol i matade satsodlingar. En sluten kolbalans för C – och P-begränsande förhållanden indikerade att inga relevanta mängder av andra (biprodukter)produkter bildades (tabell 3). Denna skillnad i biproduktspektrum kan förklara ökningen av 3-hk-utbytet per glukos.
i jämförelse med de odlingar som utförs i kemostater visar de specifika glukosupptagningshastigheterna bestämda i fed-batchkulturer betydligt högre värden på 0,75 och 0,77 mmol gCDW−1 h−1 för C – respektive P-begränsande förhållanden. Som nämnts ovan sattes börvärdet för den exponentiella faktorn för foderprofilen till 0,05 h−1 och varierade upp till 0,059 h-1, vilket resulterade i en 30-45% högre specifik tillväxttakt i fed-batch-odlingar jämfört med kemostat-experimenten med 0.04 h – 1 (Tabell 3), vilket orsakar ökat specifikt glukosupptag. För C-begränsade fermentationer var emellertid biomassautbytet på glukos jämförbart mellan kemostat och fed-batch odling. På liknande sätt höjdes de specifika 3-hk-produktionshastigheterna i matade satsodlingar, med 0,24 mmol gCDW−1 h−1 respektive 0,38 mmol gCDW−1 h−1. 3-HP-utbyten per substrat var jämförbara mellan kemostat och fed-batch odling, vilket tyder på att avkastningen är en robust odlingsparameter som är resistent mot störningar som induceras av mindre fluktuationer i odlingsuppsättningen. Inget kvarvarande fosfat, såväl som glukos, detekterades i proverna från de olika reaktorerna för P-begränsande förhållanden, vilket bekräftar att kulturerna är begränsade med avseende på motsvarande begränsning. På liknande sätt detekterades ingen kvarvarande glukos för C-begränsning i proverna.
här presenterade förbättringar av ett småskaligt kemostatsystem, som tidigare utvecklats av Klein et al. , innefattade bland annat en ökning av antalet reaktorer till 24 och en minskning av odlingsvolymen till 6,5 mL. Denna ökade grad av parallellisering gör systemet till ett lämpligt screeningverktyg med hög genomströmning, eftersom olika jämförbara småskaliga kemostatsystem arbetar med högre arbetsvolymer och med ett lägre antal reaktorer: det kontinuerliga parallella skakade bioreaktorsystemet (CosBios) använder sex respektive åtta parallella odlingsfartyg med en odlingsvolym på 20-25 mL . Det för kontinuerliga odlingar modifierade engångsbioreaktorsystemet (bioreaktor, 2mag AG) arbetar med åtta parallella reaktorer med en arbetsvolym på 10 mL . Mini-chemostat (MC) – systemet utvecklat av Bergenholm et al. omfattar 16 parallella reaktorer och kräver en arbetsvolym på 40 mL. Vårt system är därför väl tillämpligt för enkel och kostnadseffektiv screening av mikrobiell prestanda i kontinuerligt läge. Det kan dock vara av relevans för att ytterligare utvidga systemet genom att övervaka ytterligare parametrar, såsom avgaser, för att möjliggöra en detaljerad analys av kolfördelningen, vilket är särskilt viktigt för fysiologiska stamkarakteriseringar.
som vår studie föreslog att N – och P-begränsning skulle vara gynnsam för 3-HP – bildning i S. cerevisiae, skulle framtida experiment kunna innebära ytterligare screening av tillväxthastigheter med n-och P-begränsning tillämpad för att hitta den optimala 3-HP-produktionen med respektive begränsning. Vår studie visade vidare överförbarheten av fysiologiska parametrar från kemostat till matade satsodlingar. Detta anser endast vara möjligt om inga toxiska eller hämmande föreningar ackumuleras under fed-batch-odlingen, eftersom fysiologiska parametrar skulle förändras över tid och avvika från de förvärvade parametrarna i kemostater. Som ett allmänt tillvägagångssätt kan kemostatförsök fungera som ett verktyg för att undersöka påverkan av en potentiell giftig eller hämmande förening genom att tillsätta substansen till själva foderet. Eftersom alla andra parametrar är konstanta kan påverkan av föreningen och dess koncentration direkt bedömas och utvärderas. I vår studie upptäcktes ingen biproduktbildning i fed-batch-odlingar och ett förmodligen liknande stressrespons inducerades på grund av svaga syror, vilket gjorde det möjligt för fysiologiska förhållanden i småskaliga kemostater att likna dem i fed-batch-förhållanden.
Sammanfattningsvis visade denna studie att begreppet C-och P-begränsande villkor för produktion av 3-HP undersöktes i ett nytt parallelliserat kemostatodlingssystem och kunde framgångsrikt överföras till 1-L bänkbioreaktorer som arbetar i fed-batch-läge. Därför kan fysiologiska parametrar som förvärvats i kemostater användas för design och prestationsbedömning av fed-batch-odlingar med användning av avkastningsbaserade parametrar för processuppsättningen.