
introduktion
cell-eller vävnadsförnyelse och regenerering är de två huvudutvecklingskraven för vuxna organismer. Båda processernahar som utgångspunkt en population av stamceller, normalt lokaliserade i en specifik miljö som kallas” nisch”, som ger dem de nödvändiga signalerna för att upprätthålla stamnegenskaperna, eller att differentiera till de nödvändiga olika celltyperna (Figur 1). Stamcellsproliferation och differentiering måste samordnas meddöd av cellerna som behöver bytas ut. Dessutom processersåsom cellmigration, epigenetik och cellulär kommunikation är också nödvändiga för korrekt cellförnyelse . Snabb förnyelsevävnader kan erkännas av en högre mitotisk aktivitet. Omvänt innehåller långsamma förnyelsevävnader mindre mitos och kan inte lätt erkännas från icke-förnyande områden som också kan presentera somemitos . Ödet beslut av stamceller under proliferationdirekt påverka vävnadsförnyelse och homeostas. Därför är det viktigt att förstå de regleringsmekanismer som upprätthåller en balanseradcelldelning och differentiering. Extracellulära signaler (t.ex. vävnadsmikromiljö, intracellulär ROS och cytokiner) såväl som intracellulära faktorer (t. ex. epigenetiska maskiner, transkriptionsfaktorer och DNA-skadesvar) är ansvariga för reglering av stamcellsdelning.
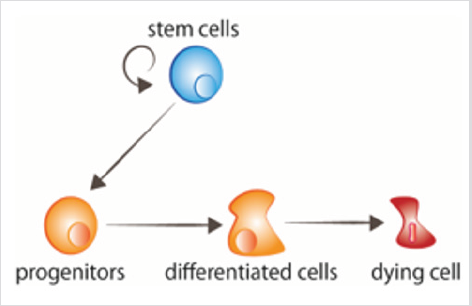
figur 1: cellomsättning stamceller proliferera, givingrise till stamfäder som därefter tar emot signalerna todifferentiate. Åldrade celler får signaler att dö.
stamceller visar tre möjliga alternativ för uppdelning: asymmetrisk uppdelning, där en stamcell och en engagerad dottercell har sitt ursprung; symmetriskt engagemang, vilket ger två engagerade dotterceller; och symmetrisk uppdelning,som ger två dotterceller som upprätthåller stamcellsegenskaper Figur 2. Även om det kan förutsägas att asymmetrisk uppdelning är den enda mekanismen som möjliggör upprätthållandet av en stabil stamcellspopulation, visade de aktuella data från lineagetracingexperiment att i de flesta vävnader uppnås balansen mellan stamcellsproliferation och generering av differentierad avkomma på nivån för hela stamcellspopulationen.Förlusten av stamceller på grund av differentiering eller cellskada inducerarsymmetrisk uppdelning för att fylla detta gap . Efter stamcellsdelning, celler som följer differentieringsprocessen passerar genom olikastadier som definieras av en kombination av transkriptionsfaktorersom styr aktiviteten hos den lämpliga repertoaren av gener, ochtillåta deras engagemang och terminaldifferentiering. För varje celllinje är slutprodukten av beslutssekvensen en specifikifferentierad celltyp figur 3a.under de flesta omständigheter är cellulär identitet – produkten av normal differentiering – stabilinom vävnader,och dess underhåll är avgörande för normal vävnadfunktion. Sådan stabilitet uppnås genom epigenetisk reglering-t .ex. Histon – demetylering och acetylering-som resulterar i arvsmönster av vävnadsspecifikt genuttryck.
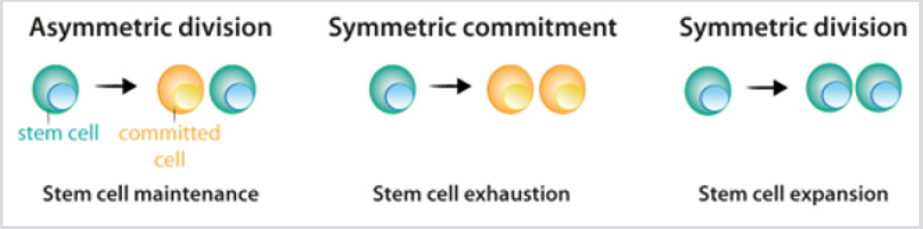
figur 2: Delningsmönster av stamceller.
a) Under asymmetrisk uppdelning ger stamceller upphov till en stamcell, som upprätthåller stamcellspopulationen och en cellsom blir engagerad i differentiering.
b) under symmetriskt engagemang ger stamcellsdelning upphov till två dotterceller som blev engagerade idifferentiering.
c) under symmetrisk uppdelning ger stamcellsdelning upphov till två stamceller. Som förklaras i texten, nuvarande experimentelldata indikerar att de tre delningssätten kan uppstå samtidigt som stamcellspopulationen upprätthålls.
emellertid kan förlust av cellidentitet uppstå. Faktum är att celler fråndrosophila imaginal skiva kan transdeterminera och förvärvaett nytt vuxen öde efter transplantation . I denna situation verkar extracellulära signaler omprogrammera någon föregångare ellerdifferentierade celler för att förvärva egenskaper hos antingen en mer stamstat eller ett nytt differentierat tillstånd. Det finns två mekanismergenom vilka en cell kan ändra sin identitet: dedifferentieringochtransdifferentiering. Dedifferentiering avser processen genomvilken en differentierad eller engagerad cell förvärvar characteristicsof en mindre mogen cellfigur 3b. Det mest dramatiska exemplet på dedifferentiering är in vitro-omvandlingen av terminallydifferentierade celler till pluripotenta celler (inducerade pluripotentstemceller, iPSCs), genom överuttryck av ett begränsat antal transkriptionsfaktorer . Transdifferentiering sker däremot när en differentierad cell ändrar det transkriptionsprogram ochkonverterar till en annan differentierad celltyp. Processen kaninträffa genom ett intermedierat steg av dedifferentiering mot aless moget stadium före omvandlingen till den nya differentierade cellen, eller direkt, utan det intermedierade steget figur 3c-d.den direkta omvandlingen av fibroblaster till myoblaster genom ektopiskuttryck av MyoD är ett exempel på den andra processen .
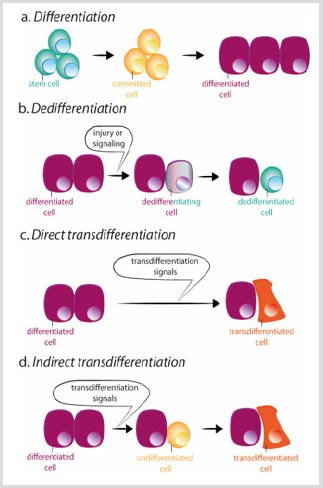
figur 3: system för differentiering, dedifferentiering och transdifferentiering.
a) under normal differentiering ger stamceller upphov till engagerade celler som i sin tur skiljer sig åt i olika celltyper.
b) dedifferentiering består av förvärv av stamcellsegenskaper av en differentierad cell. Transdifferentiering kan uppståpå ett direkt eller indirekt sätt.
c) under direkt transdifferentiering förvärvar en differentierad cell transkriptionsprogrammet av en annan celltyp, vanligtvisnära relaterad, som till exempel exokrina till endokrina pankreasceller, blir en annan differentierad cell.
d) under indirekt transdifferentiering dedifferentierar en differentierad cell innan den antar det nya transkriptionsprogrammet av den andra celltypen.
dedifferentiering och transdifferentiering förekommer också iett naturligt sätt som svar på en skada eller vävnadsförlust .Dedifferentiering förekommer till exempel naturligt under limbregeneration i urodele-amfibierna. Efter amputation av lemmar, celler intill såret dedifferentierar och bildar ett blastema sombestår av odifferentierade celler som sprider sig och så småningom omdirigeras till samma celltyp för att skapa alla komponenterav den förlorade lemmen. Naturlig transdifferentiering sker indirekt: först dedifferentierar cellen; och sedan aktiveras det naturliga utvecklingsprogrammet, vilket gör att cellen kan differentieras tillNy härstamning. Tsonis och medarbetare beskrev en naturligmekanism för transdifferentiering i en newt. De fann att närlinser avlägsnas, pigmenterade epitelceller från dorsal iristransdifferentiera och regenerera den saknade vävnaden. För att uppnådetta måste pigmenterade epitelceller först dedifferentiera ochspridas för att skapa nya linsceller och sedan differentieras tillmogna celler i linsen . I båda situationerna – dedifferentieringoch redifferentiering till samma celltyp eller transdifferentiat till en ny celltyp-kan ett komplext nätverk av signalvägar kontrollera transkriptionsprogrammet som förvärvats av varje cell i den perfekta tidspunkten.
hus, den spatiotemporala kontrollen av genuttryck krävs kontinuerligt under djurhomeostas och under aregenerativ process. Under regenerering måste emellertid celler anpassa sig till den nya situationen, vilket kräver att man fattar mergrundade beslut på cellulär nivå, ofta inklusive processer avdifferentiering och transdifferentiering som under homeostasiär knappa.
- Clevers H, Loh KM, Nusse R (2014) signalering av stamceller. Ett integralprogram för vävnadsförnyelse och regenerering Wnt-signalering och stemcell-kontroll. Vetenskap 346 (6205): 1-9.
- Leblond C P, Walker B(1956) förnyelse av cellpopulationen 36 (2): 255-276.
- Roostaee A, Benoit YD, Boudjadi S, Beaulieu JF (2016) epigenetik iintestinal Epitelcellförnyelse. J Cell Physiol 231 (11): 2361-2367.
- Ito K, Ito K (2016) Metabolism och kontroll av beslut om cellöde Ochstamcellförnyelse. Annu Rev Cell Dev Biol 32: 399-409.
- Simons BD, Clevers H (2011) strategier för homeostatiska stamceller självrenewalin vuxna vävnader. Cell 145 (6): 851-862. Dutton JR, Banga a, Slack JM W, et al. (2013) Stagespecific omprogrammering av leverceller från musembryon till en beta-cellliknande fenotyp. Mech Dev 130 (11-12): 602-612.
- Merrell AJ, Stanger BZ (2016) vuxencellplasticitet in vivo: dedifferentieringoch transdifferentiering är tillbaka i stil. Nat Rev MolCell Biol 17 (7): 413-425.
- Worley MI, Setiawan L, Hariharan IK (2012) regenerering ochöverbestämning i Drosophila imaginal-skivor. Annu Rev Genet 46: 289-310.
- Raff M (2003) vuxen Stamcellsplasticitet: faktum eller artefakt? Annu Rev CellDev Biol 19: 1-22.
- Takahashi K, Yamanaka S (2006) induktion av pluripotenta Stamcellerfrån mus embryonala och vuxna Fibroblastkulturer enligt Definieradefaktorer. Cell 126 (4): 663-676. det finns en mängd olika typer av produkter. (1988) MyoDi: ett Kärnfosfoprotein som kräver en Myc-Homologiregion för att omvandla fibroblaster till myoblaster. Vetenskap 242 (4877): 405-411.
- Jopling C, Boue S,Izpisua Belmonte JC (2011) dedifferentiering, transdifferentiering och omprogrammering: tre vägar till regenerering.Nat Rev Mol Cell Biol 12 (2): 79-89.
- Tsonis PA, Madhavan M, Tancous EE, Del Rio-Tsonis K (2004) Del Rio-Tsonis, K. en newt s öga syn på lins regenerering. Int J Dev Biol 48 (8-9): 975-980.