
en viktig del av att förstå hur dessa system fungerar för att kontrollera celldelning var upptäckten att p53 påverkar både cancer ochåldrande. Tyner et al. (2002) utarbetade agenetisk strategi hos möss för att jämföra effekterna av frånvaron av p53 eller asmaller än normalt p53-protein. De tvåomutanta transgena muslinjerna hade antingen en fullständig deletion av p53-genen (p53 -) eller en stympad form av p53 (p53m, mutant)som inte hade de första sex exonerna av p53-genen (Figur 2).
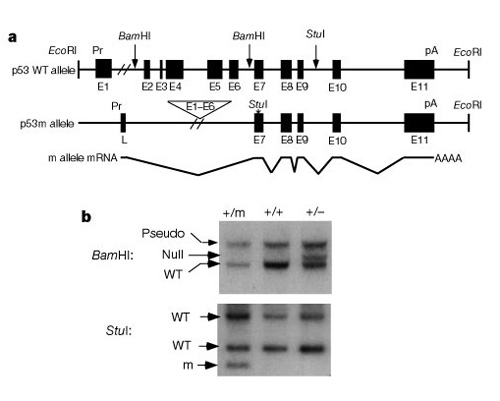
den första halvan av studien jämförde tre grupper av möss:Grupp 1, p53+/p53-(borttagningav en kopia av p53); Grupp 2, p53+/p53m(partiell deletion mutant); och Grupp 3, p53+ / p53+(vildtyp, normal). Tabell 1 visar dessa tre grupper och deras experimentella resultatrelaterade till cancer och åldrande fenotyper. Intressant nog utvecklade ingen av mössen igrupp 2, med det stympade p53-proteinet, livshotande tumörer,medan 45% av grupp 3 (vildtyp) och över 80% av grupp 1 utveckladeslivshande tumörer. Grupp 2-möss hade också en mellanliggande livslängd mellanvägenmellan den mycket korta livslängden för Grupp 1 och den längre livslängden för grupp 3wild-typmöss. Slutsatsen från dessa data är att den partiella p53-mutationen minskade förekomsten avcancer och samtidigt tycktes orsaka ett underskott i livslängden, inte en förlängd livslängd.
| Tabell 1. Experimental results from genetic mousestudies with p53 mutants | |||
| Genotype | Cancer phenotype | Agingphenotype | |
| Group 1 | p53+/p53- (complete deletion) | 80%had tumors | Muchshorter life span |
| Group 2 | p53+/p53m (partial mutant) | None | Shorterlife span |
| Group 3 | p53+/p53+ (wild type) | 45%had tumors | Normallife span |
| anpassad från Tyner etal. 2002 |
författarna ocksånoterade att Grupp 2-mössen utvecklade fenotyper som är karakteristiska för gamla möss, såsom långsam hårväxt och hunchbacked spines på grund av skelettförändringar, tidigareän de vilda mössen gjorde (figur 3).

i den andra halvan av studien, Tyner och kollegorfrågade om p53 mutantfungerade annorlunda i närvaro av normal p53. De uppfödde en ytterligare transgen muslinje med p53 – / p53m mutantalleler och fann att dessa möss inte hade det starka tumörskyddet och visadeen mycket mindre livslängd effekt. Så mutanten p53 behövde på något sätt arbeta ikonsert med normal p53 för att få effekt. Som en allmän observation rapporterade forskare att celler från p53 + / p53m heterozygot visade sig vara ungefär tre gånger så hårda totransformera som vildtypsceller. Således, även om dessa celler var resistenta motcancer, orsakade denna p53+/ p53mbakgrund också tidigare åldrande. Faktum är att aktiviteten hos p53 i dessaheterozygoter verkade vara särskilt högre än dess aktivitet i naturen type.It verkade som en sådan förändring, medan apriori, skulle vara bra mot både cancer och senescens, men det visade sig inte vara fallet.
senare, Mooreet al. visat att i odladceller med samma mutation som orsakar stympat p53-protein, kom detta stympade protein in i kärnan och colocalized med den normala p53. De studerade också p53-proteinets halveringstid i celler och fann att heterozygoterna med en kopia av mutanten p53 hade ungefär en trefaldig ökning av stabiliteten hos det normala p53-proteinet, jämfört med stabiliteten i den vilda typen ensam, vilket innebär att proteinets stabilitet förbättrades över det normala. Dessa resultat i odlade celler förlängdes och bekräftade studien utförd av Tyner et al. hos möss. Dessutom var detta den första celldelningsresponsvägen som upptäcktes för att fungera via p53 och tydligt modulera både cancerincidens och åldrande.
i RB-vägen, som kan signalera att man lämnar celldivisionscykeln, orsakar händelser som DNA-skada eller otillräcklig replikation som leder till korta telomerer vid kromosomändar minskad CDK-signalering. Detta, inturn, ökar RB-proteinkinasaktiviteten och ökar följaktligen aktiviteten hos transkriptionsfaktorn E2F. Vad är betydelsen av E2F? Denna transkriptionsfaktor binder till promotorerna av RNA-polymerasunderenheter och andra proteiner som behövs för att S-fasen ska börja, och det hjälper till att starta celldivision (Campisi 2003; Weinberg 1995). Således verkar det som att både p53 ochrb-vägarna påverkar samma cellcykelkontrollmekanismer.