
Introducere
reînnoirea și regenerarea celulelor sau țesuturilor sunt cele două cerințe principale de dezvoltare ale organismelor adulte. Ambele procese au ca punct de plecare o populație de celule stem , localizate în mod normal într-un mediu specific numit „nișă”, care le furnizează semnalele necesare pentru a menține proprietățile tulpinii sau pentru a se diferenția de diferitele tipuri de celule necesare (Figura 1). Proliferarea și diferențierea celulelor stem trebuie coordonate cumoartea celulelor care trebuie înlocuite. În plus, procesecum ar fi migrația celulară, epigenetica și comunicarea celulară,sunt, de asemenea, necesare pentru reînnoirea celulară adecvată . Reînnoire rapidățesuturile pot fi recunoscute printr-o activitate mitotică mai mare. În schimb, țesuturile cu reînnoire lentă conțin mai puțină mitoză și este posibil să nu fie ușor recunoscute din zonele care nu se reînnoiesc, care pot prezenta și unele mitoze . Deciziile soarta celulelor stem în timpul proliferăriiinfluențează direct reînnoirea țesuturilor și homeostazia. Prin urmare, înțelegerea mecanismelor de reglementare care susțin o echilibrarediviziune celulară și diferențierea este critică. Semnalele extracelulare (de exemplu, micromediul tisular, ros intracelular și citokine), precum și factorii intracelulari (de exemplu, mașinile epigenetice, factorii de transcripție și răspunsul la deteriorarea ADN-ului) sunt responsabili pentru reglementarea diviziunii celulelor stem.
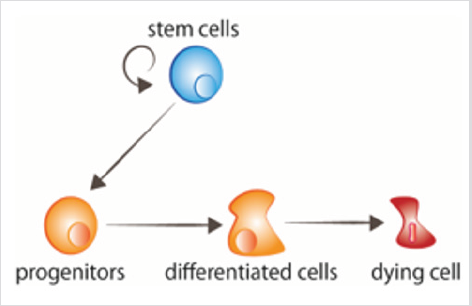
figura 1: celulele stem din turnover proliferează, dând naștere progenitorilor care ulterior primesc semnalele de diferențiere. Celulele îmbătrânite primesc semnale pentru a muri.
celulele Stem prezintă trei opțiuni posibile de divizare: diviziunea asimetrică, în care o celulă stem și o celulă fiică angajată sunt originare; angajamentul simetric, care are două celule fiice angajate; și diviziunea simetrică,care produce două celule fiice care mențin proprietățile celulelor stem Figura 2. Deși s-ar putea anticipa că diviziunea asimetrică este singurul mecanism care permite menținerea unei populații stabile de celule stem, datele actuale din experimentele liniare au demonstrat că, în majoritatea țesuturilor, echilibrul dintre proliferarea celulelor stem și generarea de surse diferențiate se realizează la nivelul întregii populații de celule stem.Pierderea celulelor stem datorată diferențierii sau deteriorării celulelor, inducediviziune simetrică pentru a umple acest gol . După diviziunea celulelor stem, celulele care urmează procesul de diferențiere trec prin diferitetape care sunt definite de o combinație de factori de transcripțiecare controlează activitatea repertoriului adecvat de gene șipermite angajamentul lor și diferențierea terminală. Pentru fiecare celulă,produsul final al secvenței de decizii este un tip de celulă specific diferit figura 3A. în majoritatea circumstanțelor, identitatea celulară – produsul diferențierii normale – este stabilăîn țesuturi, iar menținerea acesteia este crucială pentru funcționarea normală a țesutului. O astfel de stabilitate se realizează prin reglarea epigenetică-de exemplu, demetilarea histonei și acetilarea – care are ca rezultat modele ereditare ale expresiei genelor specifice țesutului .
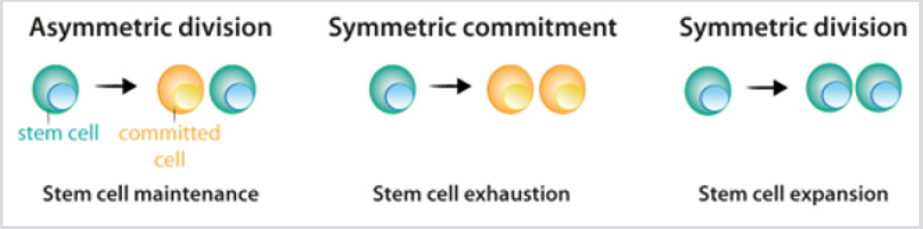
figura 2: modelul de divizare a celulelor stem.
a) în timpul diviziunii asimetrice, celulele stem dau naștere unei celule stem, care menține populația de celule stem și unei celule care se angajează să diferențieze.
b) în timpul angajamentului simetric, diviziunea celulelor stem dă naștere la două celule fiice care s-au angajat să differențieze.
c) în timpul diviziunii simetrice, diviziunea celulelor stem dă naștere la două celule stem. După cum se explică în text, experimentul curentdatele indică faptul că cele trei moduri de divizare pot apărea în timp ce se menține populația de celule stem.
cu toate acestea, poate apărea pierderea identității celulare. Într-adevăr, celulele dindrosophila imaginal disc sunt capabile să transdetermine și să dobândească o nouă soartă adultă în urma transplantului . În această situație,indicii extracelulari par să reprogrameze unele celule precursoare saucelule diferențiate pentru a dobândi caracteristici fie ale unei stări mai stem, fie ale unei noi stări diferențiate. Există două mecanismeprin care o celulă își poate schimba identitatea: dediferențierea șitransdiferențierea. Dediferențierea se referă la procesul prin care o celulă diferențiată sau angajată dobândește caracteristici ale unei celule mai puțin mature figura 3b. Cel mai dramatic exemplu de dediferențiere este conversia in vitro a celulelor terminalediferențiate în celule pluripotente (celule pluripotente induse, IPSC), prin supraexprimarea unui număr limitat de factori de transcriere . Transdiferențierea, în schimb, are loccând o celulă diferențiată o schimbă program transcripțional șise transformă într-un alt tip de celulă diferențiat. Procesul poate avea loc printr-o etapă intermediară de dediferențiere către o etapă mai puțin matură înainte de conversia în noua celulă diferențiată sau direct, fără etapa intermediară figura 3c-d.conversia directă a fibroblastelor în mioblaste prin ectopicexpresia MyoD este un exemplu al celui de-al doilea proces .
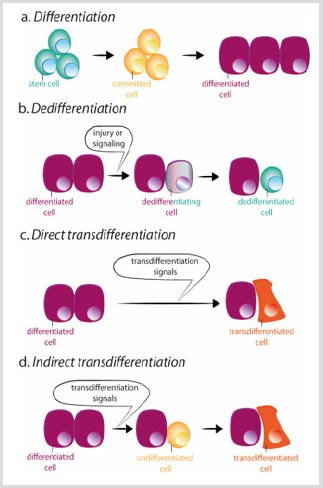
figura 3: scheme de diferențiere, dediferențiere și diferențiere trans.
a) în timpul diferențierii normale, celulele stem dau naștere unor celule angajate care, la rândul lor, se diferențiază în diferite tipuri de celule.
b) Dediferențierea constă în dobândirea proprietăților celulelor stem de către o celulă diferențiată. Se poate produce diferențierea transîn mod direct sau indirect.
c) în timpul diferențierii trans directe, o celulă diferențiată dobândește programul transcripțional al unui alt tip de celulă, de obiceiclos legate, de exemplu, exocrine la celulele pancreatice endocrine, devenind o celulă diferențiată diferită.
d) în timpul diferențierii trans indirecte, o celulă diferențiată dediferențiază înainte de a adopta noul program transcriptional al celuilalt tip de celulă.
Dediferențierea și transdiferențierea apar, de asemenea, într-un mod natural ca răspuns la o leziune sau la pierderea țesutului .Dediferențierea, de exemplu, apare în mod natural în timpul limbregenerare în amfibienii urodele. După amputarea membrelor, celulele adiacente plăgii se dediferențiază, formând un Blastem care constă din celule nediferențiate care proliferează și,în cele din urmă, se rediferențiază în același tip de celulă pentru a crea toate componenteledin membrul pierdut. Transdiferențierea naturală are loc indirect: mai întâi, celula dediferențiază; și apoi programul natural de dezvoltare este activat, permițând celulei să se diferențieze în noua linie. Tsonis și colaboratorii au descris un naturalmecanismul transdiferențierii într-un triton. Ei au descoperit că atunci cândlentile sunt îndepărtate, celulele epiteliale pigmentate din iristrans dorsal diferă și regenerează țesutul lipsă. Pentru a realiza acest lucru, celulele epiteliale pigmentate trebuie mai întâi să dediferențieze șiproliferează pentru a crea noi celule ale lentilelor și apoi să se diferențieze încelulele mature ale lentilei . În ambele situații-dediferențierea și rediferențierea în același tip de celulă sau transdiferențiat la un nou tip de celulă-o rețea complexă de căi de semnalizare poate controla programul transcripțional dobândit de fiecare celulă în momentul perfect.
hus, controlul spațiotemporal al expresiei genelor este necesar în mod continuu în timpul homeostaziei animale și în timpul procesului generativ. Cu toate acestea, în timpul regenerării, celulele trebuie să se adapteze la noua situație, ceea ce necesită luarea unor decizii mai profunde la nivel celular, adesea incluzând procese de diferențiere și transdiferențiere care în timpul homeostaziei sunt rare.
- Clevers H, Loh KM, Nusse R (2014) semnalarea celulelor Stem. Un program integrat pentru reînnoirea și regenerarea țesuturilor semnalizare Wnt și control stemcell. Știință 346 (6205): 1-9.
- Leblond C P, Walker B(1956) reînnoirea populației celulare 36 (2): 255-276.
- Roostaee a, Benoit YD, Boudjadi S, Beaulieu JF (2016) epigenetică Înînnoirea celulelor epiteliale intestinale. Celula J Physiol 231 (11): 2361-2367.
- Ito K, Ito K (2016) metabolismul și controlul deciziilor privind soarta celulelor și reînnoirea celulelor stem. Annu Rev Cell Dev Biol 32: 399-409.
- Simons bd, Clevers H (2011) strategii pentru țesuturile adulte homeostatice cu celule stem. Celula 145( 6): 851-862.
- Yang Y, Akinci E, Dutton JR, Banga A, Slack JM W, și colab. (2013) reprogramarea specifică etapelor celulelor hepatice embrionare de șoarece la un fenotip asemănător celulelor beta. Mech Dev 130 (11-12): 602-612.
- Merrell AJ, Stanger BZ (2016) plasticitatea celulelor adulte in vivo: dediferențierea și transdiferențierea sunt din nou în stil. Nat Rev MolCell Biol 17 (7): 413-425.
- Worley MI, Setiawan L, Hariharan IK (2012) regenerare și transdeterminare în discurile imaginale Drosophila. Anu Rev Genet 46: 289-310.
- Raff M (2003) plasticitatea celulelor Stem adulte: fapt sau artefact? Annu Rev Celldev Biol 19: 1-22.
- Takahashi K, Yamanaka S (2006) inducerea celulelor Stem pluripotente din culturi de fibroblaste embrionare și adulte de șoarece prin factori definiți. Celula 126( 4): 663-676.
- Tapscotr SJ, Davis RL, Thayer MJ, Cheng PF, Weintraub H, și colab. (1988) MyoDi: o fosfoproteină nucleară care solicită o regiune de omologie Myc pentru a converti fibroblastele în mioblaste. Știință 242 (4877): 405-411.
- Jopling C, Boue s, Izpisua Belmonte JC (2011) Dediferențiere,transdiferențiere și reprogramare: trei căi spre regenerare.Nat Rev Mol Cell Biol 12 (2): 79-89.
- Tsonis PA, Madhavan M, Tancous EE, Del Rio-Tsonis K (2004) Del Rio-Tsonis, K. O nouă vedere a regenerării lentilelor. Int J Dev Biol 48 (8-9): 975-980.