
proteínas de adesão celular
adesão celular é um processo selectivo, de modo a que as células adiram apenas a outras células de tipos específicos. Esta selectividade foi demonstrada pela primeira vez em estudos clássicos de desenvolvimento embrionário, que mostraram que as células de um tecido (por exemplo, fígado) aderem especificamente às células do mesmo tecido, em vez de às células de um tecido diferente (por exemplo, cérebro). Esta adesão célula-célula seletiva é mediada por proteínas transmembranares chamadas moléculas de adesão celular, que podem ser divididas em quatro grandes grupos: as selectinas, as integrinas, a superfamília imunoglobulina (Ig) (assim chamada porque eles contêm domínios estruturais semelhantes às imunoglobulinas), e as cadherinas (tabela 12.3). A adesão celular mediada pelas seletinas, integrinas e cadherinas requer Ca2+ ou Mg2+, tantas interações adesivas entre as células são Ca2+- ou Mg2+-dependentes.
quadro 12.3 moléculas de adesão celular
. as selectinas mediam interacções transitórias entre leucócitos e células endoteliais ou plaquetas sanguíneas. Existem três membros da família da selectina: a l-selectina, que é expressa em leucócitos; a e-selectina, que é expressa em células endoteliais; e a P-selectina, que é expressa em plaquetas. Como discutido anteriormente neste capítulo, as selectinas reconhecem os hidratos de carbono da superfície celular (ver Figura 12.14). Um de seus papéis críticos é iniciar as interações entre leucócitos e células endoteliais durante a migração de leucócitos da circulação para locais de inflamação dos tecidos (figura 12.62). As selectinas mediam a adesão inicial dos leucócitos às células endoteliais. Segue-se a formação de adesões mais estáveis, nas quais as integrinas na superfície dos leucócitos se ligam a moléculas de adesão intercelular (ICAMs), que são membros da superfamília Ig expressas na superfície das células endoteliais. Os leucócitos firmemente ligados são então capazes de penetrar as paredes dos capilares e entrar no tecido subjacente, migrando entre as células endoteliais.

Figura 12, 62
adesão entre leucócitos e células endoteliais. Os leucócitos deixam a circulação em locais de inflamação do tecido interagindo com as células endoteliais das paredes capilares. O primeiro passo nesta interacção é a ligação das selectinas leucocitárias (mais…)
A ligação de ICAMs para integrinas é um exemplo de um heterophilic interação, em que uma molécula de adesão na superfície de uma célula (por exemplo, um ICAM) reconhece diferentes molécula na superfície de outra célula (por exemplo, uma integrina). Outros membros da superfamília Ig mediam interações homofílicas, nas quais uma molécula de adesão na superfície de uma célula se liga à mesma molécula na superfície de outra célula. Esta ligação homofílica conduz a uma adesão selectiva entre células do mesmo tipo. Por exemplo, moléculas de adesão de células nervosas (N-CAMs) são membros da superfamília Ig expressa em células nervosas, e a ligação homofílica entre N-CAMs contribui para a formação de associações seletivas entre células nervosas durante o desenvolvimento. Existem mais de 100 membros da superfamília Ig, que mediam uma variedade de interações célula-célula.o quarto grupo de moléculas de adesão celular, as cadherinas, também apresentam especificidades de ligação homofílica. Eles não estão envolvidos apenas na adesão seletiva entre as células embrionárias, mas também são os principais responsáveis pela formação de junções estáveis entre as células nos tecidos. Por exemplo, a e-cadherina é expressa em células epiteliais, então interações homofílicas entre e-cadherinas levam à adesão seletiva de células epiteliais umas às outras. Vale ressaltar que a perda de e-cadherina pode levar ao desenvolvimento de cancros decorrentes de células epiteliais, ilustrando a importância das interações célula-célula no controle do comportamento celular. Diferentes membros da família cadherin, como N-cadherin (cadherin neural) e P-cadherin (cadherin placentária), mediam a adesão seletiva de outros tipos de células.
Cerca de vinte cadherinos clássicos diferentes, como e-cadherin, foram identificados. Além disso, uma subfamília distinta de cadherinas, chamadas protocadherinas, são expressas no sistema nervoso central, onde parecem desempenhar um papel na adesão entre neurônios em sinapses. Curiosamente, diferentes neurônios parecem expressar diferentes protocadherinas, sugerindo que as protocadherinas podem desempenhar um papel no estabelecimento de conexões específicas entre neurônios. Cerca de 50 genes de protocadherina humana foram identificados e mostraram ser organizados em três grupos de genes. Cada cluster contém vários exões de codificação, o N-terminal extracelular e transmembrana protocadherin domínios, mas apenas um único conjunto de três éxons codificação do C-terminal citoplasmático de domínio (Figura 12.63). Os clusters de genes protocadherina, portanto, parecem consistir de uma região variável, codificando múltiplos domínios extracelulares e transmembranares, ligados a uma região constante codificando um único domínio citoplásmico. Esta organização de genes de protocadherina assemelha-se, de forma notável, aos genes de imunoglobulina e receptores de células T (ver figuras 5.42 e 5.43), em que múltiplos exões de regiões variáveis são unidos a uma única região constante exon. Nos genes receptores de imunoglobulina E células T, isso ocorre como resultado de rearranjos de DNA que geram diversidade no sistema imunológico. Resta determinar se as regiões variáveis e constantes das protocaderinas são unidas no nível de DNA ou ARN (por exemplo, por splicing alternativo) e em que medida rearranjos dos genes de protocadherina podem contribuir para o estabelecimento de conexões sinápticas específicas no cérebro.
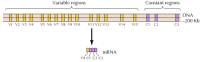
figura 12.63
Organização de grupos de genes de protocadherina. Os genes de protocadherina humana estão organizados em três grupos. No cluster ilustrado, 15 diferentes regiões variáveis que codificam domínios extracelulares e transmembranares estão ligadas a uma única região constante, (mais…)
em contraste com as junções de matriz celular estável discutidas na seção precedente, as interações célula-célula mediadas pelas selectinas, integrinas e membros da superfamília Ig são adesões transitórias em que os citoesqueletos de células adjacentes não estão ligados um ao outro. As junções de adesão estáveis envolvendo os citoesqueletos das células adjacentes são em vez disso mediadas pelas cadherinas. Como discutido no Capítulo 11, estas junções célula-célula são de dois tipos: junções adesivas e desmossomas, em que cadherinas ou proteínas relacionadas (desmogleinas e desmocolinas) estão ligadas a Feixes de actina e filamentos intermédios, respectivamente (figura 12.64). O papel das cadherinas na ligação dos citoesqueletos das células adjacentes é, portanto, análogo ao das integrinas na formação de junções estáveis entre as células e a matriz extracelular.
