
białka adhezyjne komórki
adhezja komórki-komórki jest procesem selektywnym, takim, że komórki przylegają tylko do innych komórek określonych typów. Selektywność ta została po raz pierwszy wykazana w klasycznych badaniach rozwoju zarodka, które wykazały, że komórki z jednej tkanki (np. wątroby) specyficznie przylegają do komórek tej samej tkanki, a nie do komórek innej tkanki (np. mózgu). Taka selektywna adhezja komórka-komórka jest pośredniczona przez białka transbłonowe zwane cząsteczkami adhezji komórkowej, które można podzielić na cztery główne grupy: selektyny, integryny, nadrodzinę immunoglobulin (Ig) (nazwaną tak, ponieważ zawierają domeny strukturalne podobne do immunoglobulin) i cadheriny (tabela 12.3). Adhezja komórkowa pośredniczona przez selektyny, integryny i cadheriny wymaga Ca2 + lub Mg2+, więc wiele oddziaływań adhezyjnych między komórkami zależy od Ca2+ lub Mg2+.
tabela 12.3
cząsteczki adhezyjne komórek.
selektyny pośredniczą w przemijających interakcjach między leukocytami a komórkami śródbłonka lub płytkami krwi. Istnieją trzy członkowie rodziny selektyny: l-selektyna, która ulega ekspresji na leukocytach; e-selektyna, która ulega ekspresji na komórkach śródbłonka; i P-selektyna, która ulega ekspresji na płytkach krwi. Jak omówiono wcześniej w tym rozdziale, selektyny rozpoznają węglowodany powierzchniowe komórek (patrz rysunek 12.14). Jedną z ich krytycznych ról jest inicjowanie interakcji między leukocytami a komórkami śródbłonka podczas migracji leukocytów z krążenia do miejsc zapalenia tkanek (rysunek 12.62). Selektyny pośredniczą w początkowej adhezji leukocytów do komórek śródbłonka. Po tym następuje tworzenie bardziej stabilnych zrostów, w których integryny na powierzchni leukocytów wiążą się z międzykomórkowymi cząsteczkami adhezyjnymi (iCam), które są członkami nadrodziny Ig wyrażonej na powierzchni komórek śródbłonka. Mocno przytwierdzone leukocyty są następnie w stanie przeniknąć przez ściany naczyń włosowatych i wejść do tkanki podstawowej poprzez migrację między komórkami śródbłonka.

rysunek 12.62
adhezja między leukocytami a komórkami śródbłonka. Leukocyty opuszczają krążenie w miejscach zapalenia tkanek poprzez interakcję z komórkami śródbłonka ścian naczyń włosowatych. Pierwszym etapem tej interakcji jest Wiązanie selektyn leukocytowych (więcej…)
Wiązanie iCam z integrynami jest przykładem heterofilnej interakcji, w której cząsteczka adhezji na powierzchni jednej komórki (np. ICAM) rozpoznaje inną cząsteczkę na powierzchni innej komórki (np. integryny). Inni członkowie nadrodziny Ig pośredniczą w interakcjach homofilnych, w których cząsteczka adhezyjna na powierzchni jednej komórki wiąże się z tą samą cząsteczką na powierzchni innej komórki. Takie homofilne Wiązanie prowadzi do selektywnej adhezji między komórkami tego samego typu. Na przykład cząsteczki adhezyjne komórek nerwowych (N-CAMs) są członkami nadrodziny Ig wyrażonej na komórkach nerwowych, a homofilne wiązanie między N-CAMs przyczynia się do tworzenia selektywnych skojarzeń między komórkami nerwowymi podczas rozwoju. Istnieje ponad 100 członków nadrodziny Ig, które pośredniczą w różnych interakcjach między komórkami.
czwarta grupa cząsteczek adhezyjnych komórek, cadheriny, również wykazuje homofilną specyfikę wiązania. Są one nie tylko zaangażowane w selektywną adhezję między komórkami zarodkowymi, ale są również przede wszystkim odpowiedzialne za tworzenie stabilnych połączeń między komórkami w tkankach. Na przykład, E-kadherina jest wyrażana na komórkach nabłonkowych, więc homofilne interakcje między E-kadherinami prowadzą do selektywnej adhezji komórek nabłonkowych do siebie. Warto zauważyć, że utrata e-cadherin może prowadzić do rozwoju nowotworów wynikających z komórek nabłonkowych, ilustrując znaczenie interakcji komórka-komórka w kontrolowaniu zachowania komórek. Różni członkowie rodziny cadherin, tacy jak N-cadherin (neural cadherin) I P-cadherin (placental cadherin), pośredniczą w selektywnej adhezji innych typów komórek.
zidentyfikowano około dwudziestu różnych klasycznych cadherinów, takich jak e-cadherin. Ponadto odrębna Podrodzina cadherin, zwana protokadherinami, ulega ekspresji w ośrodkowym układzie nerwowym, gdzie wydają się odgrywać rolę w adhezji między neuronami w synapsach. Intrygująco, różne neurony wydają się wyrażać różne protocadheriny, co sugeruje, że protocadheriny mogą odgrywać rolę w ustanawianiu specyficznych połączeń między neuronami. Około 50 ludzkich genów protocadherin zidentyfikowano i wykazano, że są zorganizowane w trzy klastry genów. Każdy klaster zawiera wiele eksonów kodujących N-końcowe zewnątrzkomórkowe i transmembrane protocadherin domeny, ale tylko jeden zestaw trzech eksonów kodujących C-końcowe domeny cytoplazmatyczne (rysunek 12.63). Klastry genów protocadherin wydają się składać z regionu zmiennego, kodującego wiele domen pozakomórkowych i transbłonowych, połączonych ze stałym regionem kodującym pojedynczą domenę cytoplazmatyczną. Ta organizacja genów protocadherin uderzająco przypomina geny receptorów immunoglobulin i komórek T (patrz Fig. 5.42 i 5.43), w których wiele eksonów regionu zmiennego łączy się z jednym eksonem regionu stałego. W genach receptorów immunoglobulin i komórek T zachodzi to w wyniku rearanżacji DNA, które generują różnorodność w układzie odpornościowym. Pozostaje ustalić, czy zmienne i stałe regiony protokadherin są połączone na poziomie DNA lub RNA (na przykład przez alternatywne splicing) i w jakim stopniu rearanżacje genów protokadherin mogą przyczynić się do ustanowienia specyficznych połączeń synaptycznych w mózgu.
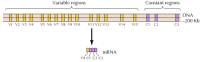
rysunek 12.63
Organizacja klastrów genów protocadherin. Ludzkie geny protocadherin są zorganizowane w trzy klastry. W klastrze ilustrowanym 15 różnych regionów zmiennych kodujących domeny zewnątrzkomórkowe i transbłonowe łączy się w jeden region stały (więcej…)
w przeciwieństwie do stabilnych połączeń komórka-matryca omówionych w poprzednim rozdziale, interakcje komórka-komórka pośredniczone przez selektyny, integryny i członków nadrodziny Ig są przejściowymi zrostami, w których cytoszkielety sąsiednich komórek nie są ze sobą połączone. Stabilne połączenia adhezyjne z udziałem cytoszkieletów sąsiednich komórek są zamiast tego pośredniczone przez cadheriny. Jak omówiono w rozdziale 11, te połączenia komórka-komórka są dwóch typów: połączenia adherenów i desmosomów, w których cadheriny lub pokrewne białka (desmogleiny i desmokoliny)są połączone odpowiednio z wiązkami aktyny i włókienkami pośrednimi (Fig. 12.64). Rola cadheryn w łączeniu cytoszkieletów sąsiednich komórek jest zatem analogiczna do roli integryn w tworzeniu stabilnych połączeń między komórkami a macierzą zewnątrzkomórkową.

Rysunek 12.
stabilne połączenia komórkowo-komórkowe za pośrednictwem cadherin. Homofilne interakcje między kadherinami pośredniczą w dwóch typach stabilnych zrostów komórkowo-komórkowych. W węzłach przylegających cadheriny są połączone z wiązkami włókien aktyny poprzez kateniny (patrz rysunek (więcej…)