
wprowadzenie
Odnowa i regeneracja komórek lub tkanek to dwa główne wymagania rozwojowe dorosłych organizmów . Oba procesy mają jako punkt wyjścia populację komórek macierzystych, Zwykle zlokalizowanych w określonym środowisku zwanym „niszą” , które dostarczają im wymaganych sygnałów do utrzymania właściwości stemness lub do różnicowania się z wymaganymi różnymi typami komórek (ryc. 1). Proliferacja i różnicowanie komórek macierzystych muszą być skoordynowane ze śmiercią komórek, które muszą zostać zastąpione. Ponadto procesy takie jak migracja komórek, epigenetyka i komunikacja komórkowa są również niezbędne do prawidłowej odnowy komórek . Szybkie odnowienie można rozpoznać po wyższej aktywności mitotycznej. Odwrotnie, powolna odnowa tkanki zawierają mniej mitozy i nie mogą być łatwo rozpoznawane z obszarów nieodnawialnych, które mogą również występować w somemitozie . Decyzje los komórek macierzystych podczas proliferacji bezpośrednio wpływają na odnowę tkanek i homeostazę. Dlatego kluczowe znaczenie ma zrozumienie mechanizmów regulacyjnych, które utrzymują zrównoważony podział i zróżnicowanie komórek. Sygnały zewnątrzkomórkowe (np. mikrośrodowisko tkanek, wewnątrzkomórkowy ROS i cytokiny) oraz czynniki wewnątrzkomórkowe (np. maszyny epigenetyczne, czynniki transkrypcyjne i odpowiedź na uszkodzenia DNA) są odpowiedzialne za regulację podziału komórek macierzystych.
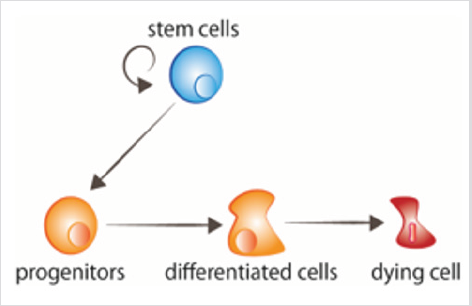
ryc. 1: rotacja komórek komórki macierzyste proliferują, dając początek komórkom progenitorowym, które następnie otrzymują sygnały różnicujące. Starzejące się komórki otrzymują sygnały do śmierci.
komórki macierzyste wykazują trzy możliwe opcje podziału: asymetryczny podział, w którym powstaje jedna komórka macierzysta i jedna komórka zależna; symetryczny podział, w którym powstają dwie komórki potomne; symetryczny podział,w którym powstają dwie komórki potomne, które zachowują właściwości komórek macierzystych ryc. 2. Chociaż można przewidzieć, że asymetryczny podział jest jedynym mechanizmem, który umożliwia utrzymanie stabilnej populacji komórek macierzystych, aktualne dane z badań lineagetracingexperiments wykazały, że w większości tkanek równowaga między proliferacją komórek macierzystych i generowaniem zróżnicowanej sprężyny jest osiągana na poziomie całej populacji komórek macierzystych.Utrata komórek macierzystych w wyniku różnicowania lub uszkodzenia komórek powoduje podziały w celu wypełnienia tej luki . Po podziale komórek macierzystych komórki, które podążają za procesem różnicowania, przechodzą przez różne etapy, które są określone przez kombinację czynników transkrypcyjnych, które kontrolują aktywność odpowiedniego repertuaru genów i ułatwiają ich zaangażowanie i końcowe różnicowanie. Dla każdej linii komórkowej produktem końcowym sekwencji decyzji jest specyficznyróżniający się Typ komórki, rysunek 3a. w większości przypadków tożsamość komórkowa-produkt normalnego różnicowania-jest stablewithin tkanek, a jej utrzymanie ma kluczowe znaczenie dla prawidłowego funkcjonowania tkanki. Taką stabilność uzyskuje się poprzez regulację epigenetyczną-np. demetylację histonów i acetylację-co skutkuje dziedziczną ekspresją genów specyficznych dla tkanki.
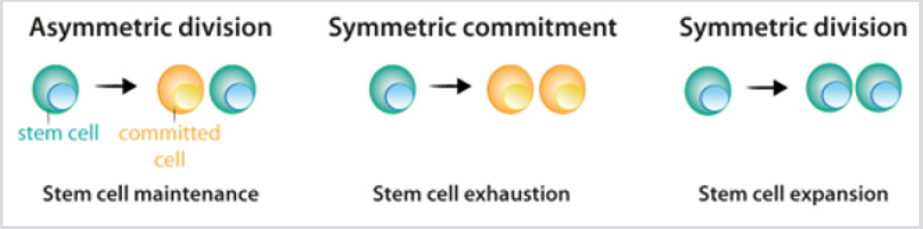
rysunek 2: Wzór podziału komórek macierzystych.
a) podczas asymetrycznego podziału komórki macierzyste dają początek jednej komórce macierzystej, która utrzymuje populację komórek macierzystych, i jednej komórce, która angażuje się w różnicowanie.
b) podczas symetrycznego podziału komórki macierzyste dają początek dwóm komórkom potomnym, które zobowiązały się do różnicowania.
c) podczas podziału symetrycznego, podział komórek macierzystych powoduje powstanie dwóch komórek macierzystych. Jak wyjaśniono w tekście, obecne eksperymentalnedata wskazuje, że trzy tryby podziału mogą wystąpić przy zachowaniu populacji komórek macierzystych.
jednak może dojść do utraty tożsamości komórki. Rzeczywiście, komórki z dysku imaginalnego Drosophila są w stanie transdeterminować i nabyć nowy los dorosłych po przeszczepieniu. W tej sytuacji pozakomórkowe sygnały zdają się przeprogramować niektóre prekursory lub różne komórki, aby uzyskać cechy bardziej stemstate lub nowego zróżnicowanego stanu. Istnieją dwa mechanizmy, dzięki którym komórka może zmienić swoją tożsamość: dedifferencjacja i transdifferencjacja. Dedifferencjacja odnosi się do procesu, w którym zróżnicowana lub zaangażowana komórka nabiera cech mniej dojrzałej komórki, rysunek 3b. Najbardziej dramatycznym przykładem dediferencjacji jest in vitro konwersja terminalnieodróżniających się komórek w komórki pluripotencjalne (indukowane komórki pluripotencjalne, iPSCs), przez nadekspresję ograniczonej liczby czynników transkrypcyjnych . Transdyferencjacja zachodzi natomiast, gdy zróżnicowana komórka zmienia swój program transkrypcyjny i przekształca się w inny zróżnicowany typ komórki. Proces może przebiegać przez pośredni etap dediferencjacji w kierunku dojrzałego etapu aless przed konwersją do nowego zróżnicowanego ogniwa, lub bezpośrednio, bez pośredniego etapu, rysunek 3c-D. bezpośrednia konwersja fibroblastów w mioblasty przez ektopicespresję miodu jest przykładem drugiego procesu .
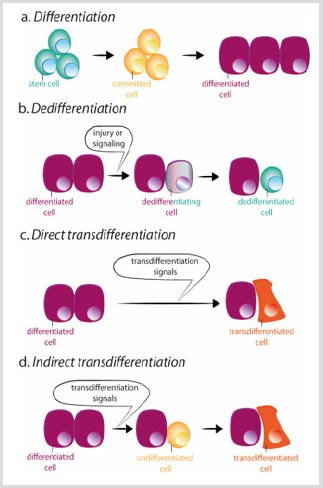
Rysunek 3: Schematy różnicowania, dedifferencjacji i różnicowania trans.
a) podczas normalnego różnicowania, komórki macierzyste dają początek komórkom zaangażowanym, które z kolei różnicują się w różnych typach komórek.
b) Dedifferencjacja polega na nabyciu właściwości komórek macierzystych przez zróżnicowaną komórkę. Różnicowanie Trans może wystąpić w sposób bezpośredni lub pośredni.
c) podczas bezpośredniego różnicowania trans, zróżnicowana komórka nabywa program transkrypcyjny innego typu komórki, Zwykle pokrewnego, jak na przykład zewnątrzwydzielnicze do endokrynologicznych komórek trzustki, stając się inną zróżnicowaną komórką.
d) podczas pośredniego różnicowania trans, zróżnicowana komórka dedyfferencjuje przed przyjęciem nowego programu transkrypcyjnego innego typu komórki.
Dedifferencjacja i transdifferencjacja występują również w naturalny sposób w odpowiedzi na uraz lub utratę tkanki .Dedifferencjacja, na przykład, występuje naturalnie podczas limbregeneracji u płazów urodele. Po amputacji kończyny, komórki przylegające do rany dedifferentiate, tworząc blastema, która składa się z niezróżnicowanych komórek, które proliferacji i ostatecznie, redifferentiate do tego samego typu komórki, aby utworzyć wszystkie składniki utraconej kończyny. Naturalna transdyferencjacja zachodzi pośrednio: najpierw dedyferencjacja komórki; a następnie aktywowany jest naturalny program rozwojowy, pozwalający komórce różnicować się w nową linię. Tsonis i współpracownicy opisali naturalny mechanizm transdyferencji w newcie. Odkryli, że po usunięciu komórek nabłonka pigmentowego z grzbietowej iristransdifferentiate i regenerują brakującą tkankę. Aby to osiągnąć, pigmentowane komórki nabłonka muszą najpierw zdedyfferencjować iproliferację, aby utworzyć nowe komórki soczewki, a następnie różnicować się w dojrzałe komórki soczewki . W obu sytuacjach-dedifferencjacjai redifferencjacja do tego samego typu komórki lub transdifferencjat do nowego typu komórki – złożona sieć szlaków sygnałowych może kontrolować program transkrypcyjny nabyty przez każdą komórkę w doskonałym punkcie czasowym.
hus, przestrzenna Kontrola ekspresji genów jest stale wymagana podczas homeostazy zwierząt oraz podczas procesu generacyjnego. Jednak podczas regeneracji komórki muszą dostosowywać się do nowej sytuacji, co wymaga podejmowania bardziej skutecznych decyzji na poziomie komórkowym, często obejmujących procesy różnicowania i transdyferencji, które podczas homeostazy są rzadkie.
- Clevers H, Loh KM, Nusse R (2014) stem cell signaling. Integralprogram do odnowy i regeneracji tkanek Wnt sygnalizacji i kontroli stemcell. Nauka 346(6205): 1-9.
- Leblond C P, Walker B (1956).
- Roostaee a, Benoit YD, Boudjadi s, Beaulieu JF (2016) epigenetyka Inintweital Epithelial cell Renewal. J Cell Physiol 231(11): 2361-2367.
- Ito K, Ito K (2016) Annu Rev Cell Dev Biol 32: 399-409.
- Simons BD, Clevers H (2011) Strategies for homeostatic stem cell selfrenewalin adult tissue. 145(6): 851-862.
- Yang y, Akinci E, Dutton JR, Banga a, Slack JM W, et al. (2013) Stagespecific reprogramming komórek wątroby zarodka myszy do komórki beta-likephenotypu. Mech Dev 130 (11-12): 602-612.
- Merrell AJ, Stanger BZ (2016) adult cell plasticity in vivo: dedifferentiationand transdifferentiation are back in style. Nat Rev MolCell Biol 17 (7): 413-425.
- Worley MI, Setiawan L, Hariharan IK (2012) Annu Rev Genet 46: 289-310.
- Raff M (2003) adult Stem Cell Plasticity: Fact or Artifact? Annu Rev CellDev Biol 19: 1-22.
- Takahashi K, Yamanaka S (2006) Induction of Pluripotent Stem Cellsfrom Mouse Embrionic and Adult Fibroblast Cultures by DefinedFactors. 126(4): 663-676.
- Tapscotr SJ, Davis RL, Thayer MJ, Cheng PF, Weintraub h, et al. (1988) MyoDi: a nuclear Phosphoprotein Reqiring a myc Homology Region toConvert Fibroblasts to Myoblasts. Nauka 242 (4877)
- Jopling C, Boue S, Izpisua Belmonte JC (2011) Dedifferentiation,transdifferentiation and reprogramming: three routes to regeneration.Nat Rev Mol Cell Biol 12 (2): 79-89.
- Tsonis PA, Madhavan M, Tancous EE, Del Rio-Tsonis K (2004) Del Rio-Tsonis, K. A newt s eye view of lens regeneration. Int J Dev Biol 48 (8-9): 975-980.