in deze studie tonen we verdere vooruitgang en toepassingsmogelijkheden van een kleinschalige continue teeltsysteem, eerder ontwikkeld door Klein et al. , voor een grotere mate van parallellisatie en een betere behandeling en controle van de afzonderlijke reactoren. De belangrijkste aspecten van de wijzigingen in het systeem zijn een verhoging van het aantal parallelle kweekvaten van 8 tot 24 reactoren en een vermindering van het werkvolume van 10 tot 6,5 mL. Het huidige systeem bestaat verder uit op maat gemaakte deksels met vier vaste poorten die worden gebruikt voor beluchting, toevoer van media, verwijdering van bouillon en inenting of bemonstering (Fig. 1). Naast de vier poorten wordt een optische staafvormige do-sonde door het deksel geplaatst voor Do-monitoring zonder het kweekproces te verstoren, en op deze manier de zuurstof fluorescerende sensorspot van de vorige opstelling te vervangen . Het waterbad, dat in de vorige opstelling een constante kweektemperatuur handhaafde, werd vervangen door een op maat gemaakt aluminium verwarmingsblok, dat aan een microplaatroerder wordt gesmolten. Aangezien de vorige versie van het kleinschalige bioreactor systeem werd gevalideerd met behulp van de splijtingsgist Schizosaccharomyces pombe , presenteren we hier de verbeterde kweek set-up voor S. cerevisiae cultivaties.
De belangrijkste operationele stappen, alsmede de aanpassing van de verdunningssnelheden door de juiste buisdiameter en pompsnelheid van de mediuminfluxpomp te kiezen, werden uitgevoerd zoals eerder beschreven . Hier werd het gewicht van het vloeistofgehalte van elke bioreactor gravimetrisch bepaald aan het einde van de kweek, zodat de respectieve verdunningssnelheid met a 5 nauwkeurig kon worden berekend.1% afwijking. Kweekbouillon en de reactorgasfase werden beide via dezelfde poort van het reactordeksel verwijderd met behulp van de effluxpomp (Fig. 1). Voor alle kweekexperimenten werden effluxpompen van 7,5 mL min−1 gebruikt. De effluxpomp was veel hoger dan de toevoerpomp, waardoor een lichte negatieve druk in het kweekvat ontstond. Dit drukverschil resulteerde in de instroom van lucht door de beluchtingspoort. De gemiddelde zuurstofmassa overdracht coëfficiënt kLa bereikt was 110 h-1, waardoor DO niveaus ruim boven 30% verzadiging tijdens het kweekproces. De pH werd niet online gecontroleerd en ook niet gecontroleerd tijdens de kweek, omdat het medium a priori werd aangepast aan een pH van 6,0, wat resulteerde in een uiteindelijke pH van 5,5 in de kweekbouillon. De pH werd dagelijks aan de lijn gemeten vanaf de uitstroom van de reactoren en na de oogst. De pH bleef constant zodra de steady-state werd bereikt en het effluent van de reactor vertoonde een geringe afwijking van 0,1 pH-eenheden (gegevens niet getoond).
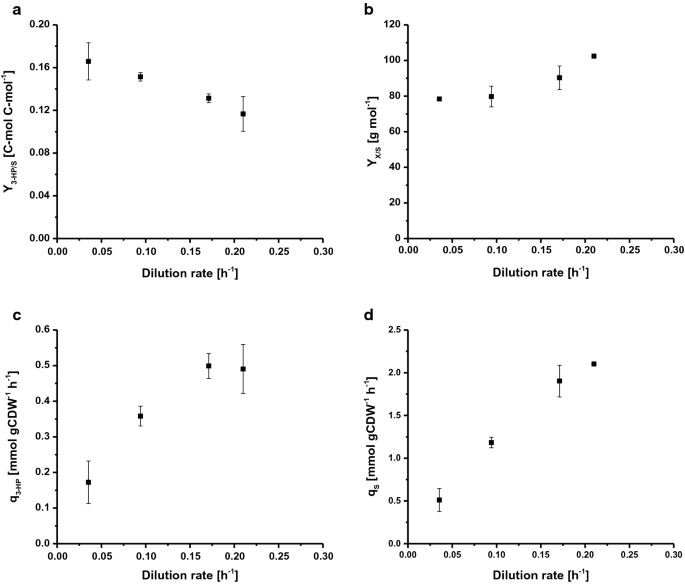
om de maximale specifieke groeisnelheid (µmax) te bepalen, werd S. cerevisiae ST938 gekweekt onder batchomstandigheden waarbij overmatige beschikbaarheid van nutriënten werd toegepast. Met behulp van glucose als koolstofbron, de µmax was 0,265 h-1, de biomassa opbrengst was 24,9 g mol−1, en de 3-PK koolstof opbrengst op glucose was 0,6% C-mol (Tabel 1). In deze teeltwijze werd het grootste deel van de koolstof omgezet in ethanol en CO2, als hoge glycolytische fluxen in wild – type S. cerevisiae zijn sterk verbonden met alcoholische fermentatie . De chemostatculturen worden gecontroleerd door de toevoer van een groeibeperkend substraat. Bij steady-state produceert wild-type S. cerevisiae geen significante hoeveelheden overloopmetabolieten onder een bepaalde groeisnelheid, wat de kritische verdunningssnelheid (Dcrit (h−1)) markeert vanwege het gebrek aan accumulatie van koolstofbron. Bij of boven deze kritische verdunningssnelheid accumuleert de koolstofbron zich in de reactor en wordt het overloopmetabolisme geactiveerd, wat resulteert in de productie van verschillende bijproducten, zoals ethanol, acetaat en kleine hoeveelheden organische zuren . Dienovereenkomstig werden continue culturen gebruikt om de relatie tussen de groeisnelheid en biomassaspecifieke 3-PK productvorming in S. cerevisiae ST938 te onderzoeken. De vier verschillende verdunningen D (h−1): 0,04, 0,09, 0,17 en 0,21 werden gekozen om onder de µmax van S. cerevisiae stam ST938 te liggen, bepaald in batchkweek. De biomassa opbrengst op glucose evenals de 3-PK opbrengst, specifieke 3-PK productiesnelheid, en de specifieke substraat opname snelheid werden onderzocht (Fig. 2a-d, Tabel 1). Alle relevante teeltparameters, d.w.z. opbrengsten en percentages met de respectieve standaardafwijkingen, zijn samengevat in Tabel 1.
Tabel 1 Teeltparameters van S. cerevisiae ST938 geteeld in continue culturen onder c-beperkte omstandigheden
Fig. 2
Selected rates and yields for C-limited aerobic chemostat cultivations of S. cerevisiae ST938 a 3-HP yield (C-mol C-mol−1) on glucose, b biomass yield on glucose (g mol−1), c specific 3-HP production rate (mmol gCDW−1 h−1) and d specific glucose uptake rate (mmol gCDW−1 h−1) at different dilution rates for S. cerevisiae ST938. Cultivations were carried out in triplicates at 30 °C and pH 5.5 under C-limited conditions. Fouten komen overeen met standaardafwijkingen afgeleid van triplicaatculturen
de teelt van S. cerevisiae ST938 onder c-limiterende omstandigheden liet een overgang zien van overwegend fermentatief metabolisme waargenomen in batchmodus naar een respiratoir metabolisme in chemostatica, wat wordt weerspiegeld in hogere opbrengsten van biomassa en in verwaarloosbare ethanol-en glycerolvorming (Tabel 1). Bovendien, kleine hoeveelheden rest glucose onder 0.In de monsters die uit de efflux van de verschillende reactoren werden genomen, werd 1 mM gedetecteerd, waarbij werd nagegaan of de culturen glucosebeperkt waren. Koolstofarme omstandigheden in steady-state chemostatculturen leken de vorming van 3-PK te bevorderen, aangezien de opbrengsten van dit product 20 tot 25 maal hoger waren dan de opbrengst bepaald in batchcultuur (Tabel 1). Dit is hoogstwaarschijnlijk toe te schrijven aan de efficiëntere omzetting van koolstof in energie in vorm van ATP tijdens ademhaling in vergelijking met fermentatie in batchculturen. Interessant is dat de cultivaties van S. cerevisiae ST938 onder c-limiterende omstandigheden toonde aan dat bij dalende specifieke groeisnelheden een constante toename van 3-PK koolstofrendementen kon worden waargenomen, met de hoogste 3-PK opbrengst van 16,6% C-mol waargenomen bij een verdunningssnelheid van 0,04 h-1 (Fig. 2 bis). Het rendement van 3 PK op biomassa is vertienvoudigd ten opzichte van het rendement in batchculturen en is meer dan verdubbeld van 0,19 tot 0,43 g gCDW−1 door de verdunningssnelheid te verlagen van 0,21 tot 0,04 h−1 (Tabel 1). De maximale specifieke productiviteit voor 3-PK van ongeveer 0,50 mmol gCDW−1 h−1 werd gezien bij verdunningssnelheden van 0,17 en 0.21 h-1, die ongeveer viermaal hoger is dan gemeten in batch Cultures (Fig. 2c, Tabel 1). Zoals verwacht nam de specifieke glucoseopnamesnelheid (QS) toe bij hogere verdunningssnelheden van 0,51 tot 2,1 mmol gCDW−1 h−1 (Fig. 2d). Deze waarden liggen tot 20 maal lager dan de maximale specifieke glucoseopnamesnelheid van 10,7 mmol gCDW−1 H−1, waargenomen bij µmax onder glucose-beperkte groeiomstandigheden in batchmodus (Tabel 1). De verkregen waarden voor de specifieke substraatopnamesnelheid en de biomassa-opbrengst bij een verdunningssnelheid van 0.09 h-1 (Tabel 1) waren in overeenstemming met gegevens van chemostatculturen met wild-type S. cerevisiae met een verdunningssnelheid van 0,10 H−1, met respectievelijk 1,1 en 1,25 mmol gCDW−1 H−1 en 0,49 g g−1 . Er is vastgesteld dat de biomassa per substraat toeneemt met toenemende verdunningssnelheden, voor wildtype S. cerevisiae, maar deze parameter is over het algemeen constant onder Dcrit zolang het onderhoudsmetabolisme een aanzienlijke koolstofput vormt die de biomassaopbrengst bij lage verdunningssnelheden vermindert . In het geval van S. cerevisiae stam ST938 bleek echter dat koolstof, dat niet gericht was op de vorming van biomassa, tot op zekere hoogte bijdroeg aan de vorming van zowel 3-PK als bijproducten.
om te concluderen, verschillen ervaren in continue culturen bij verschillende verdunningssnelheden en in vergelijking met vergelijkbare set-ups met S. cerevisiae wild-type suggereren dat de geïntegreerde biosynthetische route om 3-HP te produceren een enorme impact heeft op de gistfysiologie, die vermoedelijk wordt versterkt door adaptieve stressreacties. Ten slotte, C-beperkte chemostat teelt bij de laagste verdunningssnelheid van 0.04 h-1 resulteerde in de hoogste 3-PK koolstofopbrengst.
onderzoek naar de stikstof-en fosfaatbeperking voor verhoogde 3-HP-opbrengsten in kleinschalige chemostatculturen
na karakterisering van S. cerevisiae ST938 onder c-beperkende omstandigheden werden Cultures onder stikstof – en fosfaatbeperkende omstandigheden uitgevoerd, aangezien eerder is aangetoond dat deze gunstig zijn voor de productie van verschillende inheemse en niet – inheemse metabolieten in verschillende organismen . Hier werden de overeenkomstige Cultures onder stikstof (N) en fosfor (P) beperking onderzocht in het kleinschalige continuteeltsysteem (Fig. 1) met een ingestelde verdunningssnelheid van 0,04 h-1, die eerder had geresulteerd in de hoogste koolstofopbrengst van 3 PK onder c-beperkte omstandigheden (Tabel 1). De parameters en de respectieve standaardafwijkingen van n – en P-beperkte Cultures van giststam ST938 zijn samengevat in Tabel 2.
Tabel 2 Teeltparameters van S. cerevisiae ST938 geteeld in continue culturen met een verdunningssnelheid van 0.04 h-1 onder koolstof (C), stikstof (N) en fosfor (P) beperkte omstandigheden
voor n – en P-beperkte teeltomstandigheden werd geen residueel ammonium of fosfaat gevonden in de monsters genomen uit de efflux van de verschillende reactoren, waarbij werd gecontroleerd of de culturen beperkt waren ten opzichte van de overeenkomstige substraatbeperking (Tabel 2). Onder de N – en P-beperking werden echter kleine hoeveelheden restglucose in het bereik van 0,63 en 0,19 mM gedetecteerd (Tabel 2). Tijdens de C-beperking lagen de resterende glucosespiegels ruim onder 0,1 mM (Tabel 2). Cultivaties met n-en P-Beperkingen toonden een 3-PK opbrengst van respectievelijk 17,7 en 21,1% C-mol, met een significant hogere waarde onder p-beperkte omstandigheden in vergelijking met de 3-PK koolstof opbrengst van 16,6% C-mol berekend voor C-beperkte teeltomstandigheden (tabellen 1, 2). Evenzo werd de specifieke productiviteit voor 3-PK significant verhoogd voor n-en P-beperkte Cultures die waarden van 0,22 en 0,23 mmol gCDW−1 H−1 opleverden in vergelijking met 0,17 mmol gcdw−1 H−1 bereikt in C-beperkte culturen. Bovendien vertoonden de 3 PK-opbrengsten op biomassa vergelijkbare waarden van 0,43 en 0.44 g gCDW – 1 voor C-en N-limiterende omstandigheden werd echter een verhoogde opbrengst van 0,50 g gCDW−1 waargenomen in het geval van P-limitation. Gezien de gekwantificeerde lage concentraties bijproducten zijn deze verwaarloosbaar (Tabel 2). De waarden voor de specifieke substraatopnamesnelheid (QS) voor C – en P-beperking waren vergelijkbaar en een licht verhoogde waarde van 0,61 mmol gCDW−1 h−1 werd verkregen onder n-beperkte omstandigheden. Biomassa-opbrengsten per substraat bleken voor n-en P-beperkte omstandigheden vergelijkbare waarden van respectievelijk 72,4 en 75,2 g mol-1. Vergeleken met de biomassa opbrengst van 78.4 g mol-1 bereikt onder c-beperkte omstandigheden, deze waarden zijn vergelijkbaar. Onze studie ondersteunt de redenering dat hogere productopbrengsten per substraat werden bereikt onder n – en P-beperkende omstandigheden. Bijgevolg lijkt een hogere specifieke substraatopname in geval van n-beperking in combinatie met de minimale vorming van bijproducten uiteindelijk de productvorming te begunstigen. Onder P-beperkende omstandigheden werden de minste hoeveelheden bijproducten gevormd, wat de productsynthese zou kunnen bevorderen. Verder kan worden gespeculeerd dat sommige metabolische veranderingen mogelijk leiden tot een verlaagde hoeveelheid koolstof die als CO2 wordt vrijgegeven.
samengevat, chemostat Cultures onder n – en P-beperking toonden hogere 3-PK opbrengsten en Specifieke productiesnelheden in vergelijking met C-beperkte omstandigheden, met P-beperking die de hoogste productopbrengsten mogelijk maakte.
overdraagbaarheid van de fysiologische parameters gemeten in kleinschalige chemostatica naar fed-batchculturen in 1-L bench – top bioreactoren onder c-en P-limiterende omstandigheden
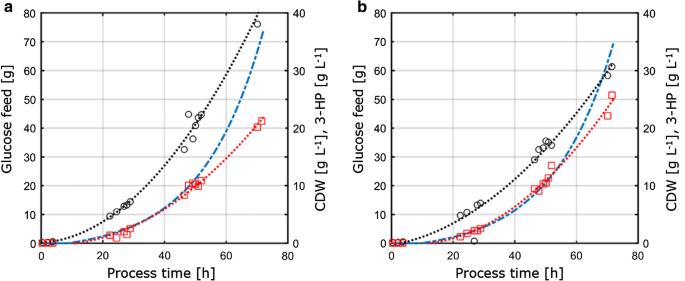
aangezien deze studie verder gericht was op de beoordeling van de overdraagbaarheid en vergelijkbaarheid van de fysiologische parameters gemeten onder verschillende kweekomstandigheden, werden de teeltparameters verkregen uit de C – en P-limiterende omstandigheden bepaald in chemostaticculturen overgebracht naar 1-L geroerde bench-top reactoren die in fed-batchmodus draaien. Het concept van deze aanpak was daarom om de belangrijkste voorwaarden en parameters constant te houden voor chemostaten en fed-batch culturen om vergelijkbaarheid te verzekeren. Deze voorwaarden omvatten (I) dezelfde C:P-verhouding als toegepast bij P-limiterende chemostatenteelt, (ii) dezelfde procesomstandigheden zoals pH en temperatuur, en (iii) dezelfde specifieke groeisnelheden als toegepast in chemostatica, bereikt via een exponentieel voedingsprofiel. Om de producttiter te optimaliseren, de parameters met de maximale productopbrengst per substraat van de chemostat experimenten (D = 0.04 h-1) onder c-en P-beperking werden gekozen voor de overdracht naar het fed-batch systeem. Als gevolg van technische beperkingen in de set-up, werden fed-batch Cultures uitgevoerd met een groeisnelheid van 0,05 h−1, wat iets hoger is in vergelijking met de ingestelde verdunningssnelheid van 0,04 h−1 in chemostat Cultures. De fed-batch teelt bestond uit een eerste batch fase om biomassa te genereren gevolgd door een exponentiële, nutriënten beperkte voedingsfase om de groeisnelheid te controleren. Alleen de voedingsfase werd relevant geacht voor de beoordeling van de overdraagbaarheid van de parameters verkregen uit het continue teeltsysteem, aangezien alleen tijdens deze fase de substraatconcentratie de microbiële groei reguleerde en dus beperkte. De fed-batch teeltprofielen van S. cerevisiae ST938 die C – en P-beperkende voorwaarden toepassen, zijn weergegeven in Fig. 3, en de overeenkomstige teeltparameters met hun respectieve standaardafwijking zijn samengevat in Tabel 3.
Fig. 3
aërobe fed-batch teelt in 1-L tafelfermentoren van S. cerevisiae ST938 met behulp van een exponentiële toevoerhelling bij d = 0,05 h-1 met het beperkende substraat a koolstof, of B fosfor. Zwarte cirkels verwijzen naar biomassatiter, rode vierkanten naar 3-PK-titer en blauwe streeplijn naar de absolute hoeveelheid glucose die naar de reactoren wordt gevoerd
Tabel 3 Teeltparameters van S. cerevisiae ST938 gekweekt in fed-batch modus in 1-L bench-top bioreactoren onder koolstof (C) en fosfor (p) limiterende omstandigheden
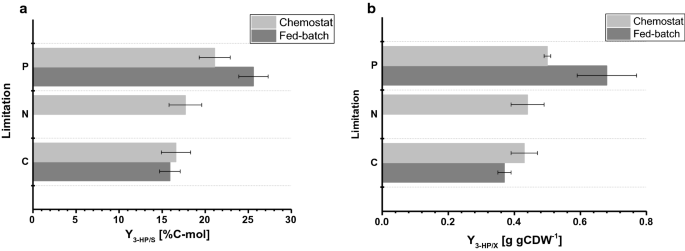
de biomassa opbrengst waargenomen in de fed-batch culturen was 77,9 en 67,7 g mol−1 voor C – en P-beperking, respectievelijk. De biomassa opbrengst voor C-beperkte Cultures was vergelijkbaar voor chemostaat en fed-batch bedrijf, terwijl voor P-beperking, een licht verminderde biomassa opbrengst werd waargenomen tijdens fed-batch bedrijf (tabellen 2 en 3), die binnen de standaardafwijkingen waargenomen voor beide biomassa opbrengsten. De koolstofopbrengst per substraat voor 3-PK was 15,9% voor C-beperking, die bijna identiek is aan de waarde berekend in het chemostatsysteem (Fig. 4a). De 3-PK opbrengsten die in deze studie werden bepaald, komen verder overeen met een koolstofopbrengst per substraat van 14% die werd verkregen in eerdere C-beperkte fed-batch studies bij pH 5,0 met een S. cerevisiae stam ontwikkeld om de β-alanine route te gebruiken en van 13% met een S. cerevisiae die de malonyl-CoA reductase-afhankelijke route gebruikt . Hogere 3-PK koolstofopbrengsten per substraat via de β-alanine route werden bepaald in studies met Escherichia coli die een waarde van 42% in fed-batch Cultures op glucose aangeven .
Fig. 4
vergelijking van kweekparameters onder verschillende beperkingen bepaald in kleinschalige chemostatica en 1-L bench-top reactoren in fed-batch modus. een koolstofrendement van 3 PK (%C-mol) en een b-rendement van 3 PK op biomassa (g gCDW-1). Fouten komen overeen met standaardafwijkingen afgeleid van drievoudige cultivaties
voor P-limiterende omstandigheden, maar onze studie toonde een aanzienlijke toename aan van 3-PK koolstofrendement van 25,6% (Fig. 4a). De waargenomen 3-PK opbrengsten op biomassa waren respectievelijk 0,38 en 0,65 g gCDW-1 voor C-en P-limiterende omstandigheden. Voor C-beperking bevestigde dit de waarden waargenomen in de chemostaatopstelling (Fig. 4b). Door de toename van de opbrengst van 3-PK per substraat onder P-limiterende omstandigheden en de lichte daling van de opbrengst van biomassa per substraat Tegelijkertijd, was de opbrengst van 3-PK per biomassa significant verhoogd in vergelijking met het chemostaatexperiment. In tegenstelling tot de cultures in chemostatica, werd geen significante accumulatie van ethanol of glycerol waargenomen in de cultures in de voeding. Uit een gesloten koolstofbalans voor C-en P-beperkende omstandigheden bleek dat er geen relevante hoeveelheden andere (bij)producten werden gevormd (Tabel 3). Dit verschil in bijproduct spectrum kan de toename van 3-PK opbrengst per glucose verklaren.
in vergelijking met de cultures uitgevoerd in chemostatica, tonen de specifieke glucoseopnamesnelheden bepaald in fed-batch culturen aanzienlijk hogere waarden van 0,75 en 0,77 mmol gCDW−1 h−1 voor respectievelijk C – en P-limiterende condities. Zoals hierboven vermeld, werd het instelpunt voor de exponentiële factor van het voerprofiel ingesteld op 0,05 h−1 en varieerde tot 0,059 h−1, resulterend in een 30-45% hogere specifieke groeisnelheid in fed-batch cultures in vergelijking met de chemostat experimenten met 0.04 h-1 (Tabel 3), wat een verhoogde specifieke glucoseopname veroorzaakt. Niettemin, voor C-beperkte fermentaties, was de biomassa opbrengst op glucose vergelijkbaar tussen chemostaat en fed-batch teelt. Evenzo werden de specifieke 3-pk productiesnelheden verhoogd in fed-batch Cultures, met respectievelijk 0,24 mmol gCDW−1 H−1 en 0,38 mmol gCDW−1 h−1. 3-PK opbrengsten per substraat waren vergelijkbaar tussen chemostaat en fed-batch teelt, wat suggereert dat opbrengsten een robuuste teeltparameter zijn die bestand is tegen verstoringen veroorzaakt door kleinere schommelingen in de teeltopstelling. In de monsters van de verschillende reactoren voor P-limiterende omstandigheden werden geen fosfaatresiduen en ook geen glucose aangetroffen, waarmee werd bevestigd dat de culturen beperkt waren ten opzichte van de overeenkomstige limiet. Ook voor C-beperking werd geen residuele glucose gedetecteerd in de monsters.
De hier gepresenteerde verbeteringen van een kleinschalig chemostatsysteem, eerder ontwikkeld door Klein et al. in het kader van dit programma werd onder meer een verhoging van het aantal reactoren tot 24 en een vermindering van het kweekvolume tot 6,5 mL opgenomen. Deze grotere mate van parallellisering maakt het systeem tot een geschikt instrument voor het screenen met hoge doorvoercapaciteit, aangezien verschillende vergelijkbare kleinschalige chemostatsystemen werken met hogere werkvolumes en met een kleiner aantal reactoren: het continuous parallel shaken bioreactor (CosBios)-systeem maakt gebruik van zes respectievelijk acht parallelle kweekvaten met een kweekvolume van 20-25 mL . Het gewijzigde bioreactor-systeem voor éénmalig gebruik (bioREACTOR, 2mag AG) werkt met acht parallelle reactoren bij een werkvolume van 10 mL . Het mini-chemostat (MC) systeem ontwikkeld door Bergenholm et al. bevat 16 parallelle reactoren en vereist een werkvolume van 40 mL. Ons systeem is daarom goed toepasbaar voor de eenvoudige en kosteneffectieve screening van microbiële prestaties in continue modus. Het zou echter van belang kunnen zijn om het systeem verder uit te breiden door aanvullende parameters te monitoren, zoals uitlaatgassen, om een gedetailleerde analyse van de koolstofdistributie mogelijk te maken, wat van bijzonder belang is voor de karakterisering van fysiologische spanningen.
aangezien onze studie suggereerde dat de N – en P-beperking gunstig zijn voor de vorming van 3-PK IN S. cerevisiae, zouden toekomstige experimenten verdere screening van groeisnelheden met n – en P-beperking kunnen inhouden om de optimale 3-P productie met elke respectievelijke beperking te vinden. Onze studie toonde verder de overdraagbaarheid van fysiologische parameters van chemostaat naar fed-batch cultivaties. Dit is alleen haalbaar als er zich tijdens de kweek van de gevoede partij geen toxische of remmende verbindingen ophopen, aangezien fysiologische parameters in de loop van de tijd zouden veranderen en zouden afwijken van de verkregen parameters in chemostatica. Als algemene benadering kunnen chemostatexperimenten dienen als hulpmiddel om de invloed van een potentieel toxische of remmende verbinding te onderzoeken door de stof toe te voegen aan het voer zelf. Aangezien alle andere parameters constant zijn, kan de invloed van de verbinding en zijn concentratie direct worden beoordeeld en geëvalueerd. In onze studie werd geen bijproductvorming gedetecteerd in fed-batch cultivaties en een vermoedelijk vergelijkbare stressreactie werd veroorzaakt door zwakke zuren, waardoor fysiologische omstandigheden in kleinschalige chemostatica konden lijken op die in fed-batch omstandigheden.
samenvattend toonde deze studie aan dat het concept van C – en P-beperkende voorwaarden voor de productie van 3-PK werd onderzocht in een nieuw parallelliseerd chemostaatteeltsysteem en met succes kon worden overgebracht naar 1-L bioreactoren die in fed-batch-modus werken. Daarom kunnen fysiologische parameters verkregen in chemostatica worden gebruikt voor het ontwerp en de prestatiebeoordeling van gevoede batchculturen met behulp van op opbrengst gebaseerde parameters voor de procesopstelling.