
celadhesie eiwitten
cel-celadhesie is een selectief proces, zodanig dat cellen zich alleen hechten aan andere cellen van specifieke typen. Deze selectiviteit werd eerst aangetoond in klassieke studies van embryoontwikkeling, die aantoonden dat cellen van één Weefsel (b.v., lever) specifiek aan cellen van hetzelfde weefsel eerder dan aan cellen van een ander weefsel (b. v., hersenen) hechten. Dergelijke selectieve cel-celadhesie wordt bemiddeld door transmembrane proteã nen genoemd celadhesie molecules, die in vier belangrijke groepen kunnen worden verdeeld: selectins, integrins, immunoglobulin (Ig) superfamilie (zo genoemd omdat zij structurele domeinen gelijkend op immunoglobulins bevatten), en cadherins (tabel 12.3). De adhesie van de cel die door selectins, integrins, en cadherins wordt bemiddeld vereist Ca2 + of Mg2+, zo zijn vele zelfklevende interactie tussen cellen Ca2 + – of Mg2 + – afhankelijk.
Tabel 12.3
Celadhesiemoleculen.
De selectinen bemiddelen voorbijgaande interacties tussen leukocyten en endotheelcellen of bloedplaatjes. Er zijn drie leden van de selectinfamilie: L-selectin, die op leukocyten wordt uitgedrukt; E-selectin, die op endotheelcellen wordt uitgedrukt; en P-selectin, die op bloedplaatjes wordt uitgedrukt. Zoals eerder in dit hoofdstuk besproken, herkennen de selectinen koolhydraten op het celoppervlak (zie figuur 12.14). Een van hun kritieke rollen is om de interactie tussen leukocyten en endotheelcellen tijdens de migratie van leukocyten van de omloop aan plaatsen van weefselontsteking in werking te stellen (figuur 12.62). Selectins bemiddelen de aanvankelijke adhesie van leukocyten aan endothelial cellen. Dit wordt gevolgd door de vorming van stabielere adhesions, waarin integrins op de oppervlakte van leukocyten aan intercellular adhesiemolecules (ICAMs) binden, die leden van de superfamilie IG zijn die op de oppervlakte van endothelial cellen worden uitgedrukt. De stevig bevestigde leukocyten kunnen dan de wanden van haarvaten doordringen en het onderliggende weefsel ingaan door tussen endothelial cellen te migreren.

figuur 12.62
adhesie tussen leukocyten en endotheelcellen. Leukocyten verlaten de omloop bij plaatsen van weefselontsteking door met de endothelial cellen van capillaire muren in wisselwerking te staan. De eerste stap in deze interactie is de binding van leukocytenselectinen (meer…)
de binding van ICAMs aan integrines is een voorbeeld van een heterofiele interactie, waarbij een adhesiemolecuul op het oppervlak van een cel (bijvoorbeeld een ICAM) een ander molecuul herkent op het oppervlak van een andere cel (bijvoorbeeld een integrine). Andere leden van de IG superfamilie bemiddelen homofiele interacties, waarbij een adhesiemolecuul op het oppervlak van een cel bindt aan hetzelfde molecuul op het oppervlak van een andere cel. Dergelijke homofiele binding leidt tot selectieve adhesie tussen cellen van hetzelfde type. De molecules van de zenuwceladhesie (N-CAMs) zijn bijvoorbeeld leden van de superfamilie IG die op zenuwcellen wordt uitgedrukt, en de homofiele band tussen N-CAMs draagt bij aan de vorming van selectieve associaties tussen zenuwcellen tijdens ontwikkeling. Er zijn meer dan 100 leden van de IG superfamilie, die een verscheidenheid van cel-celinteracties bemiddelen.
de vierde groep van celadhesiemoleculen, de cadherinen, vertonen ook homofiele bindingsspecificaties. Zij zijn niet alleen betrokken bij selectieve adhesie tussen embryonale cellen maar zijn ook primair verantwoordelijk voor de vorming van stabiele verbindingen tussen cellen in weefsels. Bijvoorbeeld, wordt e-cadherin uitgedrukt op epitheliaale cellen, zodat leiden de homofiele interactie tussen e-cadherins tot de selectieve adhesie van epitheliaale cellen aan elkaar. Het is opmerkelijk dat het verlies van e-cadherin tot de ontwikkeling van kanker kan leiden die uit epitheliaale cellen voortkomt, die het belang van cel-celinteractie in het controleren van celgedrag illustreren. De verschillende leden van de cadherinfamilie, zoals n-cadherin (neurale cadherin) en P-cadherin (placentale cadherin), bemiddelen selectieve adhesie van andere celtypes.
Ongeveer twintig verschillende klassieke cadherines, zoals E-cadherine, zijn geïdentificeerd. Bovendien komt een aparte subfamilie van cadherinen, protocadherinen genoemd, tot expressie in het centrale zenuwstelsel waar ze een rol lijken te spelen in adhesie tussen neuronen bij synapsen. Intrigerend, lijken de verschillende neuronen om verschillende protocadherins uit te drukken, die voorstellen dat protocadherins een rol in de oprichting van specifieke verbindingen tussen neuronen kunnen spelen. Ongeveer 50 menselijke protocadheringenen zijn geà dentificeerd en getoond om in drie genclusters worden georganiseerd. Elke cluster bevat veelvoudige exons die de n-eind extracellulaire en transmembrane protocadherin domeinen coderen, maar slechts één enkele reeks van drie exons die het C-eind cytoplasmic domein coderen (figuur 12.63). De protocadherin genclusters schijnen zo uit een veranderlijk gebied te bestaan, die veelvoudige extracellulaire en transmembrane domeinen coderen, verbonden met een constant gebied dat één enkel cytoplasmic domein codeert. Deze organisatie van protocadherin genen lijkt opvallend op die van immunoglobulin en T-celreceptorgenen (zie figuren 5.42 en 5.43), waarin veelvoudige veranderlijke regio exons aan één enkel constant gebied exon worden verbonden. In immunoglobulin en T-celreceptorgenen, komt dit als resultaat van herschikkingen van DNA voor die diversiteit in het immuunsysteem produceren. Het blijft te bepalen of de veranderlijke en constante gebieden van protocadherins op het niveau van DNA of RNA (bijvoorbeeld, door alternatieve splicing) worden aangesloten en in welke mate herschikkingen van protocadheringenen tot de oprichting van specifieke synaptische verbindingen in de hersenen zouden kunnen bijdragen.
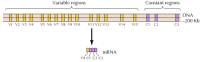
figuur 12.63
organisatie van protocadherine-genclusters. De menselijke protocadheringenen zijn georganiseerd in drie clusters. In de geà llustreerde cluster, zijn 15 verschillende veranderlijke gebieden die extracellulaire en transmembrane domeinen coderen verbonden aan één enkel constant gebied, (meer…)
in tegenstelling tot de stabiele cel-matrixverbindingen die in de vorige paragraaf werden besproken, zijn de cel-celinteracties gemedieerd door de selectines, integrines en leden van de IG-superfamilie tijdelijke adhesie waarbij de cytoskeletten van aangrenzende cellen niet met elkaar verbonden zijn. De stabiele adhesieverbindingen die cytoskeletons van aangrenzende cellen impliceren worden in plaats daarvan bemiddeld door cadherins. Zoals besproken in hoofdstuk 11, zijn deze cel-cel juncties van twee types: adherens junctions en desmosomes, waarin cadherins of verwante proteã nen (desmogleins en desmocollins) worden gekoppeld aan actin bundels en intermediaire filamenten, respectievelijk (figuur 12.64). De rol van cadherins in het verbinden van cytoskeletons van aangrenzende cellen is zo analoog aan dat van integrins in het vormen van stabiele verbindingen tussen cellen en de extracellulaire matrijs.

Figuur 12.64
stabiele cel-celverbindingen gemedieerd door de cadherinen. De homofiele interactie tussen cadherins bemiddelt twee types van stabiele cel-cel adhesie. In adherensverbindingen, zijn de cadherins verbonden met bundels van actin filamenten via de catenins (zie figuur (meer…)