
Glykolyse
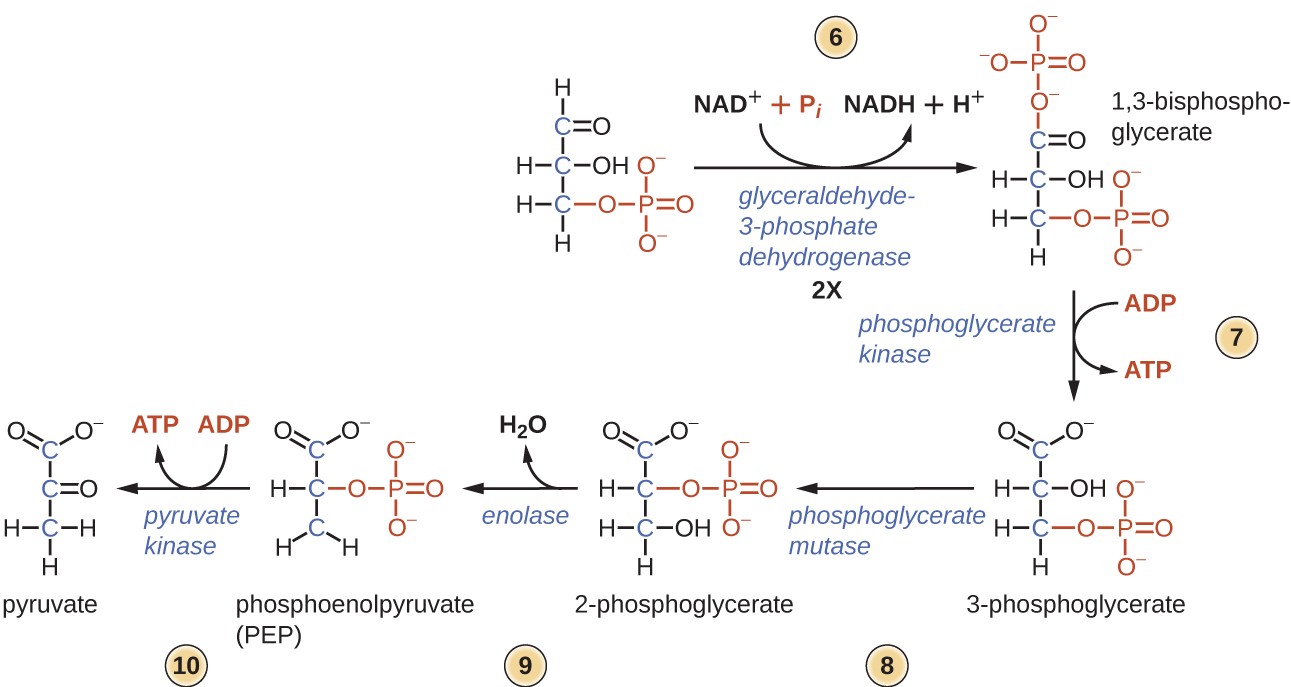
den første halvdelen av glykolyse bruker to ATP-molekyler i fosforyleringen av glukose, som deretter deles i to tre-karbonmolekyler.

Entner–Doudoroff Pathway
Entner–Doudoroff-Banen er En metabolsk vei som omdanner glukose til etanol og garn en ATP.

Pentosefosfatveien
pentosefosfatveien, også kalt fosfoglukonatveien og heksosemonofosfat shunt, er en metabolsk vei parallelt med glykolyse som genererer NADPH og fem-karbon sukker samt ribose 5-fosfat, en forløper for syntese av nukleotider fra glukose.

TCA Cycle
In this transition reaction, a multi-enzyme complex converts pyruvate into one acetyl (2C) group plus one carbon dioxide (CO2). Acetylgruppen er festet til Et Koenzym en bærer som transporterer acetylgruppen til Stedet For Krebs syklusen. I prosessen dannes et molekyl AV NADH.

i sitronsyresyklusen er acetylgruppen fra acetyl CoA festet til et firekarbonoksaloacetatmolekyl for å danne et seks-karbon citratmolekyl. Gjennom en rekke trinn oksyderes citrat, og frigjør to karbondioksidmolekyler for hver acetylgruppe matet inn i syklusen. I prosessen produseres tre NADH, en FADH2 og EN ATP eller GTP (avhengig av celletypen) ved substratnivå fosforylering. Fordi sluttproduktet av sitronsyresyklusen også er den første reaktanten, går syklusen kontinuerlig i nærvær av tilstrekkelige reaktanter. (kreditt: modifikasjon av arbeid av «Yikrazuul» / Wikimedia Commons)
Beta Oksidasjon
under fettsyreoksidasjon kan triglyserider brytes ned I 2c acetylgrupper som kan gå inn I Krebs syklusen og brukes som energikilde når glukosenivået er lavt.

Oksidativ Fosforylering
elektrontransportkjeden er en serie elektronbærere og ionpumper som brukes Til å pumpe H + – ioner over en membran. H + strømmer deretter tilbake gjennom membranen VED HJELP AV ATP-syntase, som katalyserer DANNELSEN AV ATP. Plasseringen av elektrontransportkjeden er den indre mitokondrielle matrisen i eukaryote celler og cytoplasmisk membran i prokaryote celler.

Calvin-Benson Syklus
Calvin-Benson syklusen har tre stadier. I trinn 1 inkorporerer Enzymet RuBisCO karbondioksid i et organisk molekyl, 3-PGA. I trinn 2 reduseres det organiske molekylet ved hjelp av elektroner levert AV NADPH. I trinn 3 regenereres RuBP, molekylet som starter syklusen, slik at syklusen kan fortsette. Bare ett karbondioksidmolekyl er innlemmet om gangen, så syklusen må fullføres tre ganger for å produsere et enkelt TRE-karbon GA3P molekyl, og seks ganger for å produsere et seks-karbon glukose molekyl.
