
Chaetocerotaceae, Chaetoceros:
By: Sarah Scott



Introduction:
Chaetoceros is the largest genus of marine plank-tonic diatoms. Their cells form chains that are coiled, curved, or straight. Long setae emerge from corners of the cells. それは珪藻の最大の属の一つであり、およそ400種であり、そのほとんどは海洋であり、モントレー湾で発見されたいくつかの孤立した種があった。 Chaetoceros種は世界的に分布しており、しばしば海洋生態系を支配しています。 いくつかは、海洋と沿岸の生息地の両方で重要な花を引き起こしており、いくつかの種は休息胞子および/または補助胞子を形成している。 多くの場合、異なるChaetoceros種を区別することは非常に困難であり、この大きな属を亜属に再構築するためのいくつかの試みがなされているが、研究はまだ進行中である。 Chaetoceros属は1844年にEhrenbergによって南極海の標本から最初に検出され、細胞はガードルの眺めで多かれ少なかれ長方形であり、通常弁の眺めで楕円である。 隣接する細胞の反対側のsetaeは、その起源の近くに触れ、細胞は鎖に結合し、非常に特徴的な胞子を有し、setaeの基部は短い。 Setaの構造は弁の表面のコーナーで始まる延長空プロセスである。 これらのsetaeは、一般に、末端および鎖間setaeを含み、前者は鎖の末端にあり、後者は鎖内にある。 時には、いくつかの種は、特別なintercalary setaeと呼ばれる異常なintercalary setaeを持っている、これらは一般的なintercalary setaeよりも厚く、はるかに強く珪化しています。 直径は10–50μ m、長さは6-30μ m、形状は楕円形の円筒形である。
Chaetocerosは非常に軽く珪化したfrustulesが付いている中心の珪藻である。 各錐体は、四つの長い、薄い棘、またはsetaeを持っています。 セタエは、いくつかの細胞のコロニーを形成するために一緒に錐台をリンクします。 フラスタールは通常、ガードルビューで見られます。 Chaetocerosは主に海洋の属であり、米国の内陸水域には少数の代表者しかいません。 内陸部の細胞は、生理食塩水または塩水で汚染された河川や湖沼で最大の存在量に達します。 Chaetocerosは、多くの場合、堆積物中に保存されている重く珪化静止細胞を生成します。 高い成長速度および高濃度の脂質を含むため、Chaetocerosは、生物燃料のために脂質を収穫するための潜在的な藻類として使用されてきた(Spaulding,S.およびEdlund,M.、2008)。
水柱が層状化され、水温躍層が存在する月中旬に、珪藻は一般的により支配的な植物プランクトンである。 Chaetoerosデビルは、彼らの沈没を遅く体の細胞から延びる小さな剛毛を持っています。 このChaetocerosはメイン湾の独特な植物プランクトンである。 各セルの角から、長い薄いsetaeまたは剛毛は外側に伸び、隣接するセルと融合し、この配置は長さ2mmまでの長い螺旋を作成することができます。 この生物は長いように見えるかもしれませんが、各細胞は実際には区別され、隣接する細胞から分離されています。 一緒に結合することによって、それらは表面積を増加させる。 この長い鎖に結合することは細胞が太陽からのエネルギーを捕獲する機会の多くを持っているより長い一定期間の表面に残ることを保障する。
Chaetocerosのような植物プランクトンの物理的性質は、海洋生物に物理的に損傷を与える可能性があります。 属Chaetocerosから珪質棘は、刺激を引き起こす魚の鰓フィラメントに固執することができます。 粘液は、炎症を緩和するために鰓をコートするために魚によって作成されます。 より大きな粘液産生を促進する上で、鰓はもはや水から酸素を抽出するのに十分な効率がなく、それによって魚が窒息で死ぬ原因となる。 このタイプの開花はまれですが、1991年10月にワシントン州のDabob Bayで、細胞の豊富さが1リットルあたり103細胞に達した事件がありました(Runyan、2014)。
形態:種Chaetocerosの
は、成熟中に休息している胞子の二つの90°回転が存在することを特徴とする。 まず、母細胞の頂端面での回転により、休止胞子の弁面が母細胞の弁面に平行から垂直に回転し、母細胞の弁面で第二の90°回転により、休止胞子の弁面が狭いガードル面から母細胞の広いガードル面に回転する。 これは、Chaetocerosの成熟中に休眠胞子の二つの90°回転の最初の報告です。 (Yang Li、Suying Zhu、Nina Lundholm、Songhui Lu、2015)。 Chaetoceros tenuissimusの一般的な形態は、Hyalochaete亜属のほとんどのメンバーの形態と非常によく似ています。 この亜属はGran(1897)によって定義され、最近ではEvensen&Hasle(1975)によって改訂された。 それは、細胞の体内(setaeではない)にのみ、薄い付属器および葉緑体を有する繊細な形態の鎖または孤立した細胞のいずれかを含む。 新しく湧出した水におけるchaetoceros属のような無傷の珪藻休息胞子の有病率は,湧出したシステムを播種するために混合および現在のパターンを利用する胞子の能力を示している。 より高い沈降速度は胞子形成の重要な利点であると思われ、栄養枯渇後の無愛想な環境から個体群を急速に除去し、沖合の移流を防止し、個体群を湧出の中心に近い状態に維持する。
形態は中心である:h5>
-
細胞はrapheシステムを欠いており、有意な運動性を欠いています
-
細胞はfultoportulae(struttedプロセス)とrimoportulae(labiateプロセス)を持っている可能性があります
-
有性生殖はoogamousです
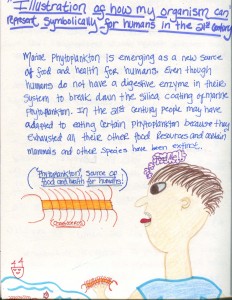
私の生物は21世紀の人間にとって象徴的に表すことができると思います食糧源。 海洋植物プランクトンは、人間のための食糧と健康の新しい供給源として浮上しています。 人間は海洋植物プランクトンのシリカコーティングを分解するための消化酵素をシステムに持っていないにもかかわらず。 21世紀には、他のすべての食糧資源を使い果たし、特定の哺乳類や他の種が絶滅したため、人々は特定の植物プランクトンに適応した可能性があります。 h5>
細胞はrapheシステムを欠いており、有意な運動性を欠いています
細胞はfultoportulae(struttedプロセス)とrimoportulae(labiateプロセス)を持っている可能性があります
有性生殖はoogamousです
科学的分類:
ドメイン:真核生物
王国:Chromalveolata
門:Heterokontophyta
クラス:Bacillariophyceae
h5>亜目:biddulphiineae
家族:cheatocerotaceae
属: 私の生物Chaetocerosのアニメーション。

私の生物Chaetocerosのアニメーション。