
細胞の密度は何ですか?
リーダーモード
生物学的材料の密度は、私たちの実験室のチューブとマルチウェルプレートの底に細胞を沈降させる責任があり、非常に多くの生物学者の日常生活の一部であるルーチンの遠心分離の基礎として機能します。 細胞とその水っぽい外観との間の密度のこれらの非常に同じ違いは、位相顕微鏡画像で観察されるコントラストの基礎でもあります。 これらの相違は実験室の設定の外でまた重要である。 例えば、プランクトンは、黒くなった深さに沈むのではなく、日光が豊富な海の深さにとどまるために、この密度差と競合しなければならない。 ほとんどの生物学者や生化学者が密度に基づく分離を日常的に利用していることを考えると、表1に収集された密度が実際にどのように議論され
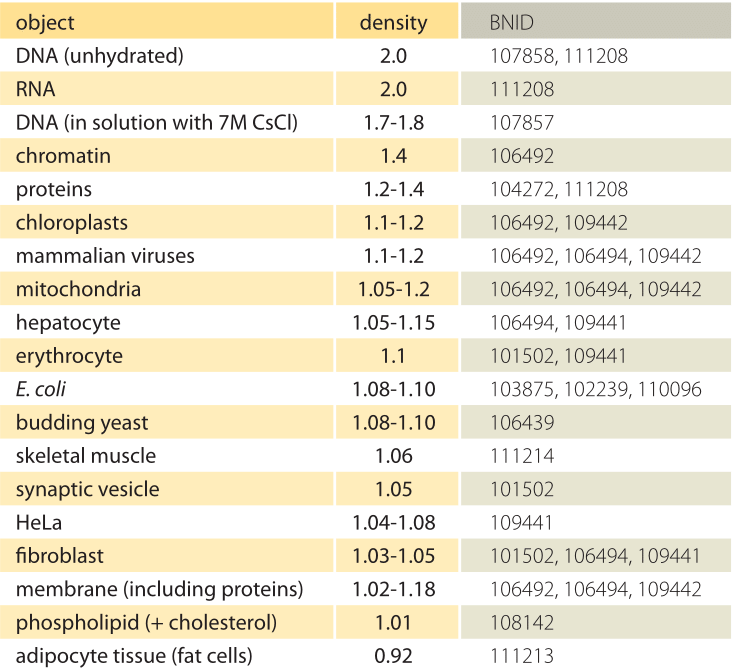
表1:水に対する生物学的オブジェクトの密度。 これは、g/mlまたは1000kg/m3の単位でそれらを与えることとほぼ同等です。 値は降順に並べ替えられます。 特に記載のない限り、値は、スクロースまたはフィコール溶液中で測定した。異なる細胞小器官および細胞型の様々な密度の基礎となる基礎は何ですか?
異なる細胞小器官および細胞型の様々な密度の基礎となる基礎は何ですか?
大部分は、これらの違いは、含水量と乾燥質量との比に起因する可能性がある。 タンパク質は、水に対して≤1.3-1.4(BNID104272、101502)の密度を有する(またはg/mlまたは1000kg/m3の単位でほぼ同等である)。 水の密度のための1のベンチマーク値を考えると、1と1の間のセル密度のための中間値のスペクトル。3は、タンパク質および水の相対的な存在量に基づいて得られる。 脂質は、約1(BNID1 0 8 1 4 2)の密度で水の隣の低端にある。 他の極端では、≤1.5(BNID103206)の密度と≤1.7のヌクレオチドを持つデンプン顆粒は、全体の質量バランスを反対方向にシフトさせることができます。
密度を知ることは、多くの場合、ショ糖またはDNA、塩化セシウムの場合に生成される濃度の勾配を含む遠心分離機で紡糸したときに、特定の生物学的成分が沈降する位置に基づいている。 密度は体積で割った質量を反映しますが、溶液中の荷電化合物の密度は、いわゆる結合水の殻によっても影響されます。 この場合の密度は有効密度となり、結合した水によって減少し、したがって塩濃度に幾分依存する(BNID107858)。
遠心分離機で発生する沈降速度は、70S、23Sなどの名前の起源であるSvedbergの単位で定量化されます。 リボソームとそのrRNAサブユニットのために。 23S rRNAは、通常の重力下で23×10-12m/sの速度で沈降する。 百万gの加速を生じる超遠心では、速度は比例して23×10-6m/sまたは約1mm/分にスケールされる。 沈降速度は、分子の密度、サイズおよび形状に依存する。 同様の形状および密度の場合、沈降速度は分子量の平方根としてスケールする。 このような場合、分子量は沈降速度の二乗となり、リボソームの23Sおよび16Sサブユニットは、おおよそ(23/16)2または約2の比率を持つ分子量を有し、0.9および0の測定値と密接にインライン化されている。それぞれ5MDa(BNID110972、110967)。 診療所では、赤血球(赤血球)の沈降速度が炎症を測定するために日常的に使用されています。 10mm/時間よりもはるかに高い速度は、通常、炎症状態の一般的な指標である沈降促進因子フィブリノーゲンの存在を示す。
水は細胞の最も豊富な分子画分であることはよく知られていますが、正確にはどれくらい豊富ですか? 多細胞生物の組織を調べると、水分含量を見つけることは、乾燥前後の組織の質量を測定する簡単な作業です。 しかし、どのようにして細胞のためにそのような測定を行うことができますか? 乾燥前と乾燥後の細胞の質量を測定するとき、周囲に水がない細胞だけをどのように測定するのですか? 遠心分離の後でさえも乾燥した固まり自体についての曖昧さに終って細胞の餌に残っている水がある。
再び放射性同位体標識が救助に来る(Cayley et al1991)。 まず、標識された水(トリチウム、3Hを使用)をセルペレット中で測定する。 これは、細胞の内側と外側の水の合計を示します。 次いで、1 4C−イヌリンまたは3H−PEGのような、標識されているが細胞に入ることができない別の可溶性化合物は、遠心分離されたペレット(例えば、e.coliでは、ペレット体積の約2 5〜3 5%(BNID))中の細胞外の水の体積を測定する。 違いは、細胞内の水分含有量を示します。 このような方法は、出芽酵母および赤血球のための質量で≧60-65%から大腸菌およびアメーバD.discoideumのための≧70%、ラット筋肉およびブタ心臓組織のための≧80%までの典型的な値をもたらす(BNID105938、103689)。 乾物の寄与は密度≤1.3(すなわちタンパク質)の成分によって支配されているので、これは≤1.1(BNID103875、106439、101502)の特徴的な全体的な密度につながる。 これらの特徴的な画分から、体積当たりの乾燥質量は、約3 0 0〜5 0 0mg/ml(BNID1 0 8 1 3 1、1 0 8 1 3 5、1 0 8 1 3 6)であると推測することができるが、成長が遅い間の値はより高くなり得る。 従って低い密度は周囲水よりより少しの密度の浮力の部分がある水中植物および乾燥した種で共通で、浮かぶようにそれらがする。 水の密度よりも低い密度は、ケルプやいくつかの細菌のようにガスによって、または周囲の媒体よりも低い分子量(MW)の溶質を使用することによって達成することができる(例えば、ナトリウムをMW≤23でアンモニウムをMW≤18で置き換える)。
人間は約60%の水(細胞で40%、間質液で15%、血漿で5%、BNID110743)で作られており、私たちのほとんどはほんの数杯でも飲むのを忘れた後に脱水の強い効果を経験 しかし、いくつかの細胞は、その含水量の減少に対して驚くほど堅牢であり得る。 例えば、ラット肝細胞におけるグルコース代謝の速度は、細胞内水分の25%の損失によって影響されなかった。 このような減少は、細胞外液の浸透性(溶質濃度)を変化させることによって達成することができる。 極端な例は、顕著な塩水エビのことです。 外の塩濃度が変動し、非常に高い環境で生活していると、不可逆的な損傷なしにわずか2%の水に乾燥させることができ、37%以上の水和レベル(完全に水和された状態の約半分)でそれらの生理学が正常に動作する嚢胞を有することが示された。 水の損失に直面したこの堅牢性は、セル内部の水の二つの形態の間で時々行われる区別に関連している可能性があります。 通常の”バルク水”は、より不必要であり、細胞成分に関連し、適切な機能のために不可欠である溶媒として役立つ”結合水”である。

 iv id=”
iv id=”div