
細胞接着タンパク質
細胞-細胞接着は、細胞が特定のタイプの他の細胞にのみ接着するような選択的 この選択性は、ある組織(例えば、肝臓)からの細胞が、異なる組織(例えば、脳)の細胞ではなく、同じ組織の細胞に特異的に接着することを示した胚発生の古典的研究で最初に実証された。 このような選択的細胞-細胞接着は、細胞接着分子と呼ばれる膜貫通タンパク質によって媒介され、セレクチン、インテグリン、免疫グロブリン(Ig)スーパーファミリー(免疫グロブリンに類似した構造ドメインを含むことから命名された)、およびカドヘリン(表12.3)に分けることができる。 セレクチン、インテグリン、およびカドヘリンによって媒介される細胞接着は、Ca2+またはMg2+を必要とするので、細胞間の多くの接着相互作用は、Ca2+
表12.3
細胞接着分子。 セレクチンは、白血球と内皮細胞または血小板との間の一過性の相互作用を媒介する。
セレクチンは、白血球と内皮細胞または血小板との間の一過性の相互作用を媒介する。 セレクチンファミリーには、白血球上で発現されるL-セレクチン、内皮細胞上で発現されるE-セレクチン、血小板上で発現されるP-セレクチンの三つのメンバーがある。 この章で前述したように、セレクチンは細胞表面炭水化物を認識します(図12.14を参照)。 それらの重要な役割の1つは、循環から組織炎症部位への白血球の移動中に白血球と内皮細胞との間の相互作用を開始することである(図12.62)。 セレクチンは、内皮細胞への白血球の初期接着を媒介する。 これに続いて、白血球の表面上のインテグリンが内皮細胞の表面上に発現されたIgスーパーファミリーのメンバーである細胞間接着分子(ICAMs)に結合するより安定 しっかりと付着した白血球は、その後、毛細血管の壁を貫通し、内皮細胞間を移動することによって下にある組織に入ることができる。

図12.62
白血球と内皮細胞との間の接着。 白血球は、毛細血管壁の内皮細胞と相互作用することによって組織炎症の部位で循環を残す。 この相互作用の最初のステップは、白血球セレクチンの結合である(詳細は、白血球セレクチンの結合を参照)。..インテグリンへのICAMsの結合は、ある細胞(例えば、ICAM)の表面上の接着分子が別の細胞(例えば、インテグリン)の表面上の異なる分子を認識するヘテロ親和性 Igスーパーファミリーの他のメンバーは、1つの細胞の表面上の接着分子が別の細胞の表面上の同じ分子に結合するホモフィリック相互作用を媒介する。 このような相親和性結合は、同じタイプの細胞間の選択的接着をもたらす。 例えば、神経細胞接着分子(N−Cam)は、神経細胞上に発現されるIgスーパーファミリーのメンバーであり、N−Cam間の同族性結合は、発達中の神経細胞間の選択的会合の形成に寄与する。 Igスーパーファミリーには100人以上のメンバーがおり、様々な細胞-細胞相互作用を媒介している。
細胞接着分子の第四のグループ、カドヘリンは、また、ホモフィリック結合特異性を表示します。 それらは胚細胞間の選択的接着に関与するだけでなく、組織中の細胞間の安定な接合部の形成にも主に関与している。 例えば、e-カドヘリンは上皮細胞上で発現されるので、E-カドヘリン間の同族性相互作用は、上皮細胞の相互への選択的接着をもたらす。 これは、e-カドヘリンの損失は、細胞の挙動を制御する細胞-細胞相互作用の重要性を示す、上皮細胞から生じる癌の発症につながることができること N-カドヘリン(神経カドヘリン)およびP-カドヘリン(胎盤カドヘリン)などのカドヘリンファミリーの異なるメンバーは、他の細胞型の選択的接着を媒介する。
E-カドヘリンのような約二十種類の古典的なカドヘリンが同定されている。 さらに、protocadherinsと呼ばれるcadherinsの明瞭なサブファミリーはシナプスでニューロン間の付着の役割を担うようである中枢神経系で表現されます。 興味深いことに、異なるニューロンが異なるプロトカドヘリンを発現するように見え、プロトカドヘリンがニューロン間の特定の接続の確立に役割を果 約50個のヒトプロカドヘリン遺伝子が同定され、三つの遺伝子クラスターに組織されることが示されている。 各クラスターには、N末端の細胞外および膜貫通プロトカドヘリンドメインをコードする複数のエクソンが含まれていますが、C末端の細胞質ドメインをコードする3つのエクソンの1つのセットのみが含まれています(図12.63)。 プロトカドヘリン遺伝子クラスターは,単一の細胞質ドメインをコードする定常領域にリンクされた複数の細胞外および膜貫通ドメインをコードする可変領域からなるように見える。 プロトカドヘリン遺伝子のこの組織は、複数の可変領域エクソンが単一の定常領域エクソンに結合されている免疫グロブリンおよびT細胞受容体遺伝子(図5.42および5.43参照)のそれに著しく類似している。 免疫グロブリンおよびT細胞受容体遺伝子において、これは、免疫系において多様性を生成するDNA再編成の結果として生じる。 プロトカドヘリンの可変領域と定常領域がDNAまたはRNAレベルで結合されているかどうか(例えば、代替スプライシングによって)、プロトカドヘリン遺伝子のどの程度の再配列が脳内の特定のシナプス接続の確立に寄与するかどうかを決定する必要がある。
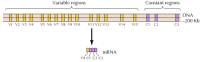
図12.63
プロトカドヘリン遺伝子クラスターの組織。 ヒトのプロトカドヘリン遺伝子は三つのクラスターに組織されている。 例示されるクラスターでは、細胞外および膜貫通ドメインをコードする1 5の異なる可変領域が、単一の定常領域に連結されている(詳細は、1 9 9 9年、1 9 9 9年、1 9 9 9年、1 9 9 9..)
前のセクションで説明した安定な細胞-マトリックス接合とは対照的に、セレクチン、インテグリン、およびIgスーパーファミリーのメンバーによって媒介される細胞-細胞相互作用は、隣接する細胞の細胞骨格が互いにリンクされていない一過性の癒着である。 隣接する細胞の細胞骨格を含む安定した接着接合は、代わりにカドヘリンによって媒介される。 第11章で説明したように、これらの細胞-細胞接合は、カドヘリンまたは関連タンパク質(デスモグレインおよびデスモコリン)がそれぞれアクチン束および中間フィラメントに連結されているアドヘレン接合およびデスモソームの二つのタイプである(図12.64)。 隣接する細胞の細胞骨格を連結するカドヘリンの役割は、細胞と細胞外マトリックスとの間の安定な接合部を形成するインテグリンのそれに類似している。

図12.64
カドヘリンによって媒介される安定な細胞-細胞接合。 カドヘリン間のホモフィリック相互作用は、安定した細胞-細胞癒着の二つのタイプを仲介します。 Adherens接合では、カドヘリンはカテニンを介してアクチンフィラメントの束にリンクされている(図を参照してください(詳細。..p>
div