
Proteine di adesione cellulare
L’adesione cellula-cellula è un processo selettivo, tale che le cellule aderiscono solo ad altre cellule di tipi specifici. Questa selettività è stata dimostrata per la prima volta in studi classici sullo sviluppo embrionale, che hanno dimostrato che le cellule di un tessuto (ad esempio, fegato) aderiscono specificamente alle cellule dello stesso tessuto piuttosto che alle cellule di un tessuto diverso (ad esempio, cervello). Tale adesione selettiva cellula-cellula è mediata da proteine transmembrana chiamate molecole di adesione cellulare, che possono essere suddivise in quattro gruppi principali: le selectine, le integrine, la superfamiglia delle immunoglobuline (Ig) (così chiamata perché contengono domini strutturali simili alle immunoglobuline) e le cadherine (Tabella 12.3). L’adesione cellulare mediata dalle selectine, integrine e cadherine richiede Ca2 + o Mg2+, quindi molte interazioni adesive tra le cellule sono Ca2+o Mg2 + dipendenti.
Tabella 12.3
Molecole di adesione cellulare.
Le selectine mediano interazioni transitorie tra leucociti e cellule endoteliali o piastrine del sangue. Ci sono tre membri della famiglia selectina: L-selectina, che è espressa sui leucociti; E-selectina, che è espressa sulle cellule endoteliali; e P-selectina, che è espressa sulle piastrine. Come discusso in precedenza in questo capitolo, le selectine riconoscono i carboidrati della superficie cellulare (vedi Figura 12.14). Uno dei loro ruoli critici è quello di avviare le interazioni tra leucociti e cellule endoteliali durante la migrazione dei leucociti dalla circolazione ai siti di infiammazione dei tessuti (Figura 12.62). Le selectine mediano l’adesione iniziale dei leucociti alle cellule endoteliali. Questo è seguito dalla formazione di aderenze più stabili, in cui le integrine sulla superficie dei leucociti si legano alle molecole di adesione intercellulare (ICAM), che sono membri della superfamiglia Ig espressa sulla superficie delle cellule endoteliali. I leucociti saldamente attaccati sono quindi in grado di penetrare le pareti dei capillari e di entrare nel tessuto sottostante migrando tra le cellule endoteliali.

Figura 12.62
Adesione tra leucociti e cellule endoteliali. I leucociti lasciano la circolazione nei siti di infiammazione dei tessuti interagendo con le cellule endoteliali delle pareti dei capillari. Il primo passo in questa interazione è il legame delle selectine dei leucociti (altro…)
Il legame di ICAMs alle integrine è un esempio di interazione eterofila, in cui una molecola di adesione sulla superficie di una cellula (ad esempio, un ICAM) riconosce una molecola diversa sulla superficie di un’altra cellula (ad esempio, un’integrina). Altri membri della superfamiglia Ig mediano le interazioni omofile, in cui una molecola di adesione sulla superficie di una cellula si lega alla stessa molecola sulla superficie di un’altra cellula. Tale legame omofilo porta all’adesione selettiva tra cellule dello stesso tipo. Ad esempio, le molecole di adesione delle cellule nervose (N-CAMs) sono membri della superfamiglia Ig espressa sulle cellule nervose e il legame omofilo tra N-CAMs contribuisce alla formazione di associazioni selettive tra le cellule nervose durante lo sviluppo. Ci sono più di 100 membri della superfamiglia Ig, che mediano una varietà di interazioni cellula-cellula.
Il quarto gruppo di molecole di adesione cellulare, le cadherine, mostra anche specificità di legame omofilo. Non solo sono coinvolti nell’adesione selettiva tra le cellule embrionali, ma sono anche i principali responsabili della formazione di giunzioni stabili tra le cellule nei tessuti. Ad esempio, l’E-caderina è espressa sulle cellule epiteliali, quindi le interazioni omofile tra le E-caderine portano all’adesione selettiva delle cellule epiteliali l’una all’altra. È interessante notare che la perdita di E-caderina può portare allo sviluppo di tumori derivanti dalle cellule epiteliali, illustrando l’importanza delle interazioni cellula-cellula nel controllo del comportamento cellulare. Diversi membri della famiglia caderina, come N-caderina (caderina neurale) e P-caderina (caderina placentare), mediano l’adesione selettiva di altri tipi di cellule.
Sono stati identificati circa venti diversi cadherin classici, come E-cadherin. Inoltre, una sottofamiglia distinta di cadherine, chiamate protocadherine, sono espresse nel sistema nervoso centrale dove sembrano svolgere un ruolo nell’adesione tra i neuroni alle sinapsi. Curiosamente, diversi neuroni sembrano esprimere diverse protocadherine, suggerendo che le protocadherine possono svolgere un ruolo nella creazione di connessioni specifiche tra i neuroni. Circa 50 geni umani della protocaderina sono stati identificati e indicati per essere organizzati in tre gruppi genici. Ogni cluster contiene più esoni che codificano i domini extracellulari e transmembrana della protocaderina N-terminale, ma solo un singolo insieme di tre esoni che codificano il dominio citoplasmatico C-terminale (Figura 12.63). I cluster di geni della protocaderina sembrano quindi costituiti da una regione variabile, che codifica più domini extracellulari e transmembrana, collegati a una regione costante che codifica un singolo dominio citoplasmatico. Questa organizzazione dei geni della protocaderina assomiglia sorprendentemente a quella dei geni del recettore delle immunoglobuline e delle cellule T (vedi Figure 5.42 e 5.43), in cui più esoni a regione variabile sono uniti a un singolo esone a regione costante. Nei geni del recettore delle immunoglobuline e delle cellule T, ciò si verifica a seguito di riarrangiamenti del DNA che generano diversità nel sistema immunitario. Resta da determinare se le regioni variabili e costanti delle protocadherine sono unite a livello di DNA o RNA (ad esempio, mediante splicing alternativo) e in che misura i riarrangiamenti dei geni della protocadherina potrebbero contribuire alla creazione di specifiche connessioni sinaptiche nel cervello.
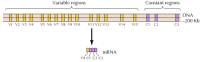
Figura 12.63
Organizzazione dei cluster di geni della protocaderina. I geni della protocaderina umana sono organizzati in tre gruppi. Nel cluster illustrato, 15 diverse regioni variabili che codificano domini extracellulari e transmembrana sono collegate a una singola regione costante, (altro…)
In contrasto con le giunzioni stabili cellula-matrice discusse nella sezione precedente, le interazioni cellula-cellula mediate dalle selectine, dalle integrine e dai membri della superfamiglia Ig sono aderenze transitorie in cui i citoscheletri delle cellule adiacenti non sono collegati tra loro. Giunzioni di adesione stabili che coinvolgono i citoscheletri delle cellule adiacenti sono invece mediate dalle caderine. Come discusso nel Capitolo 11, queste giunzioni cellula-cellula sono di due tipi: giunzioni aderenti e desmosomi, in cui cadherine o proteine correlate (desmogleine e desmocolline) sono collegate rispettivamente a fasci di actina e filamenti intermedi (Figura 12.64). Il ruolo delle cadherine nel collegare i citoscheletri delle cellule adiacenti è quindi analogo a quello delle integrine nel formare giunzioni stabili tra le cellule e la matrice extracellulare.

Figura 12.64
Giunzioni cellule-cellule stabili mediate dalle caderine. Le interazioni omofile tra cadherine mediano due tipi di aderenze cellule-cellule stabili. Nelle giunzioni adherens, le cadherine sono collegate a fasci di filamenti di actina tramite le catenine (vedi Figura (altro…)