
mi a sejtek sűrűsége?
olvasó mód
a biológiai anyag sűrűsége felelős a sejtek laboratóriumi csöveink és több kútlemezeink aljára történő leülepedéséért, és a rutin centrifugálás alapjául szolgál, amely sok biológus mindennapi életének része. Ugyanezek a sűrűségbeli különbségek a sejtek és vizes külsejük között szintén az alapja annak a kontrasztnak, amelyet a fázismikroszkópos képeken megfigyelünk. Ezek a különbségek a laboratóriumi beállításokon kívül is fontosak. Például a planktonnak meg kell küzdenie ezzel a sűrűségkülönbséggel, hogy az óceán mélyén maradjon, ahol a napfény bőséges, ahelyett, hogy a megfeketedett mélységbe süllyedne. Tekintettel arra, hogy a legtöbb biológus és biokémikus napi rendszerességgel használja a sűrűség alapján történő elválasztást, meglepőnek tűnik, hogy az 1.táblázatban összegyűjtött sűrűségeket valójában milyen ritkán tárgyalják.
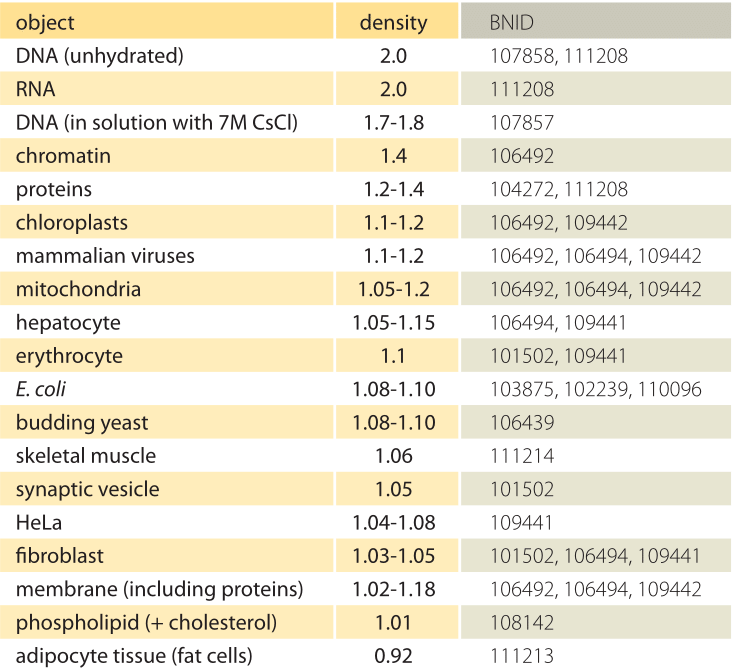
1.táblázat: a biológiai tárgyak sűrűsége a vízhez viszonyítva. Ez majdnem megegyezik azzal, hogy g/ml vagy 1000 kg/m3 egységekben adják meg őket. Az értékek csökkenő sorrendben vannak rendezve. Eltérő rendelkezés hiányában az értékeket szacharózban vagy ficoll oldatban mértük.
mi az alapja a különböző organellák és sejttípusok eltérő sűrűségének? Ezek a különbségek nagyrészt a víztartalom és a száraz tömeg arányának tulajdoníthatók. A fehérjék sűrűsége 1,3-1,4 (BNID 104272, 101502) a vízhez viszonyítva (vagy majdnem ekvivalens g/ml vagy 1000 kg/m egységben3). Tekintettel a víz sűrűségére vonatkozó 1 referenciaértékre, az 1 és 1 közötti cellasűrűség közbenső értékeinek spektruma.A 3-at a fehérjék és a víz relatív bősége alapján kapjuk meg. A lipidek az alsó végén vannak a víz mellett, körülbelül 1 sűrűségű (BNID 108142). A másik végletben a keményítőgranulátumok sűrűsége 1,5 (bnid 103206), a nukleotidok pedig 1,7-nél az ellenkező irányba tolhatják a teljes tömegmérleget.
a sűrűség ismerete gyakran azon a helyen alapul, ahol egy adott biológiai komponens leülepszik, amikor centrifugában forgatják, amely gyakran szacharóz, vagy DNS esetén cézium-klorid által termelt koncentrációk gradiensét tartalmazza. A sűrűség tükrözi a tömeget osztva a térfogattal, de az oldatban töltött vegyületek esetében a sűrűséget az úgynevezett kötött víz héja is befolyásolja. A sűrűség ebben az esetben effektív sűrűséggé válik, amelyet a kötött víz csökkent, és így némileg függ a sókoncentrációtól (BNID 107858).
a centrifugában előforduló ülepedési sebességet Svedberg egységekben számszerűsítik, amely a 70s, 23S stb. a riboszóma és annak rRNS alegységei. A 23S rRNS normál gravitáció mellett 23 60-12 m/s sebességgel üledezik. Egy millió g gyorsulást eredményező ultracentrifugában a sebesség arányosan 23-ra változik 6-6 m/s vagy körülbelül 1 mm/perc. Az ülepedés sebessége a molekula sűrűségétől, méretétől és alakjától függ. Hasonló alakzatok és sűrűségek esetén az ülepedési sebesség a molekulatömeg négyzetgyökeként skálázódik. Ilyen esetekben a molekulatömeg az ülepedési sebesség négyzetének felel meg, így a riboszóma 23S és 16S alegységeinek molekulatömege nagyjából (23/16)2 vagy körülbelül 2, amely szorosan összhangban van a 0,9 és 0 mérésekkel.5 MDa (BNID 110972, 110967). A klinikán az eritrociták (vörösvértestek) ülepedési sebességét rutinszerűen használják a gyulladás mérésére. A 10 mm/óránál jóval magasabb arányok általában a pro-ülepedési faktor fibrinogén jelenlétét jelzik, amely a gyulladásos állapot általános mutatója.
köztudott, hogy a víz a sejtek leggyakoribb molekuláris frakciója, de mennyire bőséges pontosan? Ha többsejtű organizmusokból származó szöveteket vizsgálunk, a víztartalom megtalálása egyszerű feladat a szövet tömegének mérése szárítás előtt és után. De hogyan lehet ilyen méréseket végezni a sejtek számára? Amikor szárítás előtt és után megmérjük a sejtek tömegét, hogyan mérjük csak azokat a sejteket, amelyek körül nincs víz? A centrifugálás után is víz marad a sejtpelletben, ami kétértelműséget eredményez a száraz tömeggel kapcsolatban.
ismét a rádió-izotópos címkézés jön a mentésre (Cayley et al 1991). Először a címkézett vizet (trícium felhasználásával, 3H) egy sejtpelletben mérjük. Ez jelzi a víz mennyiségét a sejteken belül és kívül. Ezután egy másik oldható vegyület, amely címkézett, de nem tud bejutni a sejtbe, például 14C-inulin vagy 3h-PEG, centrifugált pelletben méri a sejteken kívüli vízmennyiséget (például E. coliban a pellet térfogatának körülbelül 25-35% – a (BNID)). A különbség a sejtek víztartalmát jelzi. Ezek a módszerek A kezdő élesztő és a vörösvérsejtek esetében 60-65 tömegszázalékos, az E. coli és az amőba D. discoideum esetében 70% – os, a patkány izom-és sertés szívszövetei esetében pedig 80% – os (BNID 105938, 103689) tipikus értékeket eredményeznek. Mivel a szárazanyag-hozzájárulást az 1,3 sűrűségű összetevők (azaz a fehérjék) dominálják, ez a jellemző teljes sűrűséghez vezet 6,1 (bnid 103875, 106439, 101502). Ezekből a jellemző frakciókból a térfogatra jutó száraz tömeg körülbelül 300-500 mg/ml (BNID 108131, 108135, 108136) lehet, de lassú növekedés esetén az értékek magasabbak lehetnek. Az alacsony sűrűség gyakori a száraz magokban és a víz alatti növényekben, amelyek úszóképes részei kisebb sűrűséggel rendelkeznek, mint a környező víz, így lehetővé téve számukra az úszást. A vízénél alacsonyabb sűrűség vagy gázzal, mint a tengeri moszatban és néhány baktériumban, vagy a környező közegnél alacsonyabb molekulatömegű oldott anyagok (MW) alkalmazásával érhető el (pl. a nátrium helyettesítése MW-val) 23 ammóniummal MW-val 68), mint a kis rákféléknél, Antarktiszi kopepodák.
az emberek körülbelül 60% vízből állnak (40% a sejtekben, 15% az intersticiális folyadékban és 5% a vérplazmában, BNID 110743), és a legtöbbünk megtapasztalta a kiszáradás erős hatásait, miután elfelejtette inni még néhány poharat is. Mégis, egyes sejtek meglepően robusztusak lehetnek a víztartalom csökkenése szempontjából. Például a patkány májsejtekben a glükóz metabolizmus sebességét nem befolyásolta az intracelluláris víz 25% – os vesztesége. Ilyen csökkenés érhető el ozmózissal-az extracelluláris folyadék tonicitásának (oldott anyag koncentrációjának) megváltoztatásával. Szélsőséges példa a figyelemre méltó sós garnélarák. Olyan környezetben élve, ahol a külső sókoncentráció ingadozhat és nagyon magas lehet, kimutatták, hogy cisztái csak 2% vízre száríthatók visszafordíthatatlan károsodás nélkül, és 37% – nál magasabb hidratációs szintnél (a teljesen hidratált állapotnak csak körülbelül a fele) fiziológiájuk normálisan viselkedik. Ez a robusztusság a vízveszteséggel szemben összefüggésben lehet azzal, hogy néha különbséget tesznek a sejt belsejében lévő víz két formája között. Normál “ömlesztett víz”, amelyek nélkülözhetőbbek, és” kötött víz”, amely a sejtkomponensekhez kapcsolódik, és oldószerként szolgál, amely elengedhetetlen a megfelelő működéshez.




