
Sejtadhéziós fehérjék
a sejt-sejt adhézió szelektív folyamat, oly módon, hogy a sejtek csak bizonyos típusú más sejtekhez tapadnak. Ezt a szelektivitást először az embriófejlődés klasszikus tanulmányaiban mutatták ki, amelyek azt mutatták, hogy az egyik szövetből (például májból) származó sejtek specifikusan ugyanazon szövet sejtjeihez tapadnak, nem pedig egy másik szövet sejtjeihez (például agy). Az ilyen szelektív sejt-sejt adhéziót transzmembrán fehérjék, úgynevezett sejtadhéziós molekulák közvetítik, amelyek négy fő csoportra oszthatók: a szelektinek, az integrinek, az immunglobulin (ig) szupercsalád (azért nevezték el, mert az immunglobulinokhoz hasonló szerkezeti doméneket tartalmaznak), valamint a kadherinek (12.3.táblázat). A szelektinek, integrinek és kadherinek által közvetített sejtadhézió Ca2+ – t vagy Mg2+-t igényel, így a sejtek közötti sok adhéziós kölcsönhatás Ca2+ – vagy Mg2+ – függő.
12.táblázat.3
Sejtadhéziós molekulák.
a szelektinek átmeneti kölcsönhatásokat közvetítenek a leukociták és az endothel sejtek vagy a vérlemezkék között. A szelektin családnak három tagja van: az L-szelektin, amely leukocitákon expresszálódik; az e-szelektin, amely endothel sejteken expresszálódik; és a P-szelektin, amely a vérlemezkéken expresszálódik. Amint azt korábban ebben a fejezetben tárgyaltuk, a szelektinek felismerik a sejtfelszíni szénhidrátokat (lásd a 12.14.ábrát). Az egyik kritikus szerepük a leukociták és az endothel sejtek közötti kölcsönhatások megindítása a leukociták keringésből a szövetgyulladás helyére történő migrációja során (12.62 ábra). A szelektinek közvetítik a leukociták kezdeti tapadását az endothel sejtekhez. Ezt követi a stabilabb adhéziók kialakulása, amelyekben a leukociták felületén lévő integrinek kötődnek az intercelluláris adhéziós molekulákhoz (iCam-ok), amelyek az IG szupercsalád tagjai, amelyek az endoteliális sejtek felületén expresszálódnak. A szorosan kötődő leukociták ezután képesek behatolni a kapillárisok falába, és az endoteliális sejtek közötti migrációval bejutnak az alatta lévő szövetbe.

12.62 ábra
adhézió a leukociták és az endothel sejtek között. A leukociták a szövetgyulladás helyén hagyják el a keringést, kölcsönhatásba lépve a kapilláris falak endoteliális sejtjeivel. Ennek az interakciónak az első lépése a leukocita szelektinek kötődése (több…)
az ICAMs integrinekhez való kötődése példa a heterofil kölcsönhatás, amelyben egy adhéziós molekula az egyik sejt felületén (például egy ICAM) felismer egy másik molekulát egy másik sejt felületén (például egy integrin). Az Ig szupercsalád többi tagja közvetíti a homofil kölcsönhatásokat, amelyekben az egyik sejt felületén lévő adhéziós molekula ugyanahhoz a molekulához kötődik egy másik sejt felületén. Az ilyen homofil kötés szelektív tapadást eredményez az azonos típusú sejtek között. Például az idegsejt-adhéziós molekulák (N-bütykök) az idegsejteken expresszált Ig szupercsalád tagjai, az N-bütykök közötti homofil kötés pedig hozzájárul az idegsejtek közötti szelektív asszociációk kialakulásához a fejlődés során. Az Ig szupercsaládnak több mint 100 tagja van, amelyek különféle sejt-sejt kölcsönhatásokat közvetítenek.
a sejtadhéziós molekulák negyedik csoportja, a kadherinek szintén homofil kötési sajátosságokat mutatnak. Nemcsak az embrionális sejtek közötti szelektív tapadásban vesznek részt, hanem elsősorban a szövetek sejtjei közötti stabil csomópontok kialakulásáért is felelősek. Például az e-kadherin hámsejteken expresszálódik, így az e-kadherinek közötti homofil kölcsönhatások a hámsejtek szelektív tapadásához vezetnek. Figyelemre méltó, hogy az e-kadherin elvesztése a hámsejtekből származó rák kialakulásához vezethet, szemléltetve a sejt-sejt kölcsönhatások fontosságát a sejtek viselkedésének szabályozásában. A cadherin család különböző tagjai, mint például az N-cadherin (neurális cadherin) és a P-cadherin (placenta cadherin), más sejttípusok szelektív adhézióját közvetítik.
körülbelül húsz különböző klasszikus kadherint, például E-kadherint azonosítottak. Ezenkívül a cadherinek különálló alcsaládja, az úgynevezett protocadherinek, a központi idegrendszerben expresszálódnak, ahol úgy tűnik, hogy szerepet játszanak a szinapszisok neuronjai közötti tapadásban. Érdekes módon úgy tűnik, hogy a különböző neuronok különböző protocadherineket expresszálnak, ami arra utal, hogy a protocadherinek szerepet játszhatnak az idegsejtek közötti specifikus kapcsolatok kialakításában. Körülbelül 50 humán protocadherin gént azonosítottak, és kimutatták, hogy három géncsoportba szerveződnek. Minden klaszter több exont tartalmaz, amelyek az N-terminális extracelluláris és transzmembrán protocadherin doméneket kódolják, de csak egy három exonból álló készlet, amely a C-terminális citoplazmatikus domént kódolja (12.63.ábra). A protocadherin géncsoportok tehát úgy tűnik, hogy egy változó régióból állnak, amely több extracelluláris és transzmembrán domént kódol, és egy állandó régióhoz kapcsolódik, amely egyetlen citoplazmatikus domént kódol. Ez a protocadherin gének szervezete feltűnően hasonlít az immunglobulin és a T-sejt receptor génekéhez (lásd az 5.42 és 5.43 ábrákat), amelyekben több változó régió exon kapcsolódik egyetlen állandó régió exonhoz. Az immunglobulin és a T-sejt receptor génekben ez a DNS-átrendeződések eredményeként következik be, amelyek az immunrendszer sokféleségét generálják. Még meg kell határozni, hogy a protocadherinek változó és állandó régiói a DNS vagy az RNS szintjén kapcsolódnak-e egymáshoz (például alternatív splicing útján), és hogy a protocadherin gének átrendeződése milyen mértékben járulhat hozzá a specifikus szinaptikus kapcsolatok kialakulásához az agyban.
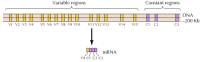
12.63 ábra
protocadherin géncsoportok szervezése. Az emberi protocadherin gének három klaszterbe szerveződnek. Az illusztrált klaszterben az extracelluláris és transzmembrán doméneket kódoló 15 különböző változó régió egyetlen állandó régióhoz kapcsolódik (több…)
az előző szakaszban tárgyalt stabil sejt-mátrix csomópontokkal ellentétben a szelektinek, integrinek és az Ig szupercsalád tagjai által közvetített sejt-sejt kölcsönhatások átmeneti adhéziók, amelyekben a szomszédos sejtek citoszkeletonjai nem kapcsolódnak egymáshoz. A szomszédos sejtek citoszkeletonjait érintő stabil adhéziós csomópontokat ehelyett a kadherinek közvetítik. Amint azt a 11. fejezet tárgyalja, ezek a sejt-sejt csomópontok kétféle típusúak: adherens csomópontok és dezmoszómák, amelyekben a kadherinek vagy rokon fehérjék (dezmogleinek és dezmokollinok) aktin kötegekhez és köztes szálakhoz kapcsolódnak (12.64 ábra). A kadherinek szerepe a szomszédos sejtek citoszkeletonjainak összekapcsolásában Analóg az integrinekével a sejtek és az extracelluláris mátrix közötti stabil csomópontok kialakításában.

12.ábra.64
a kadherinek által közvetített stabil sejt-sejt csomópontok. A kadherinek közötti homofil kölcsönhatások kétféle stabil sejt-sejt adhéziót közvetítenek. Az adherens csomópontokban a kadherinek a katenineken keresztül aktinszálak kötegeihez kapcsolódnak (Lásd az ábrát (tovább…)