
pienimuotoisen jatkuvan viljelyn järjestelmän parantaminen
tässä tutkimuksessa osoitamme Kleinin et al: n aiemmin kehittämän pienimuotoisen jatkuvan viljelyn edistysaskeleita ja käyttömahdollisuuksia. , yksittäisten reaktoreiden parallelizaation ja käsittelyn sekä valvonnan lisäämiseksi. Järjestelmään tehtyjen muutosten keskeisiä näkökohtia ovat rinnakkaisten viljelyastioiden määrän lisääminen 8 reaktorista 24 reaktoriin ja työskentelytilavuuden pienentäminen 10: stä 6,5 millilitraan. Nykyinen järjestelmä koostuu lisäksi mittatilaustyönä tehdyistä kansista, joissa on neljä kiinteää porttia, joita käytetään ilmastukseen, väliaineiden toimittamiseen, liemenpoistoon sekä inokulointiin tai näytteenottoon (Kuva. 1). Neljän portin lisäksi kannen läpi työnnetään do-tarkkailua varten optinen sauvamainen DO-anturi häiritsemättä viljelyprosessia, ja näin korvataan edellisen asetelman happi-fluoresoiva anturipiste . Vesihaude, joka edellisessä asetelmassa piti yllä tasaista viljelylämpötilaa, korvattiin mittatilaustyönä valmistetulla alumiinilämmityslohkolla, joka sulatetaan mikroliittisekoittimeen. Koska pienimuotoisen bioreaktorijärjestelmän edellinen versio validoitiin fissiohiivalla Schizosaccharomyces pombe, esittelemme tässä parannetun viljelyjärjestelmän S. cerevisiae-viljelmille.
toiminnan perusvaiheet sekä laimennusnopeuksien säätäminen valitsemalla sopiva putken halkaisija ja väliaineen virtauspumpun pumppunopeus suoritettiin edellä kuvatulla tavalla . Tässä kunkin bioreaktorin nestepitoisuuden paino määritettiin gravimetrisesti viljelyn lopussa, mikä mahdollisti vastaavan laimennusnopeuden tarkan laskemisen arvolla 5.1% poikkeama. Viljelmäliemi ja reaktorin kaasufaasi poistettiin molemmat reaktorin kannen saman portin kautta efflux-pumpulla (Kuva. 1). Kaikissa viljelykokeissa käytettiin 7, 5 mL min−1: n poistopumppunopeutta. Poistopumppunopeus oli huomattavasti suurempi kuin syöttöpumppunopeus, mikä aiheutti pienen alipaineen viljelyastian sisällä. Tämä paine-ero johti ilman virtaamiseen ilmastusportin läpi. Keskimääräinen hapenmassan siirtokerroin kLa saavutettiin 110 h-1, mikä mahdollisti DO: n reilusti yli 30 prosentin kylläisyyden koko viljelyprosessin ajan. PH: ta ei seurattu verkossa eikä kontrolloitu viljelyn aikana, sillä kasvualustan pH oli a priori säädetty 6,0: aan, jolloin lopullinen pH oli viljelyliemessä 5,5. PH mitattiin päivittäin reaktoreiden ulosvirtauksesta ja sadonkorjuun jälkeen. PH pysyi vakiona heti vakaan tilan saavuttamisen jälkeen ja reaktorin poistovedessä oli pieni poikkeama 0,1 pH-yksikköä (tietoja ei näy).
tutkittaessa 3 hv: n tuotantoa pienimuotoisissa kemostaateissa eri laimennusnopeuksilla C-rajoitusolosuhteissa
suurimman spesifisen kasvunopeuden (µmax) määrittämiseksi S. cerevisiae ST938: AA viljeltiin eräolosuhteissa, joissa käytettiin ravinteiden liiallista saatavuutta. Käyttämällä glukoosia hiililähteenä µmax oli 0,265 h−1, biomassan tuotto oli 24,9 g mol-1 ja 3-HP hiilen saanto glukoosista oli 0,6% C-mol (Taulukko 1). Tässä viljelytavassa suurin osa hiilestä metaboloitui etanoliksi ja CO2: ksi , korkeina glykolyyttisinä virtauksina villissä s-tyypissä. cerevisiae liittyy vahvasti alkoholikäymiseen . Kemostaattiviljelyä ohjataan kasvua rajoittavan kasvualustan tarjonnalla. Vakaassa tilassa luonnonvarainen S. cerevisiae-tyyppi ei tuota merkittäviä määriä ylivuodon metaboliitteja alle tietyn kasvunopeuden, mikä merkitsee kriittistä laimennusnopeutta (Dcrit (h−1)), koska hiililähde ei kasaudu. Tällä kriittisellä laimennusnopeudella tai sen yläpuolella hiililähde kerääntyy reaktoriin ja ylivuodon aineenvaihdunta käynnistyy, jolloin syntyy erilaisia sivutuotteita, kuten etanolia, asetaattia ja pieniä määriä orgaanisia happoja . Näin ollen jatkuvia viljelmiä käytettiin S. cerevisiae ST938: n kasvunopeuden ja biomassaspesifisen 3-HP: n tuotteenmuodostuksen välisen suhteen tutkimiseen. Neljän eri laimennusnopeuden D (h−1): 0, 04, 0, 09, 0, 17 ja 0, 21 valittiin alittamaan S. cerevisiae-kannan st938 µmax eräviljelyssä määritettynä. Biomassan saantoa glukoosista sekä 3 hv: n saantoa, spesifistä 3 hv: n tuotantonopeutta ja spesifistä substraatin kertymänopeutta tutkittiin (Kuva. 2a-D, Taulukko 1). Taulukossa 1 on yhteenveto kaikista merkityksellisistä viljelyparametreista eli sadoista ja määristä sekä vastaavista keskihajonnoista.
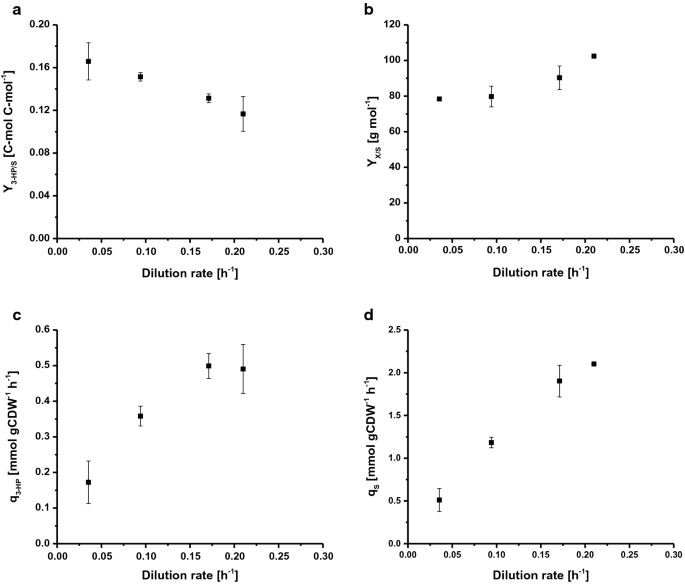
Selected rates and yields for C-limited aerobic chemostat cultivations of S. cerevisiae ST938 a 3-HP yield (C-mol C-mol−1) on glucose, b biomass yield on glucose (g mol−1), c specific 3-HP production rate (mmol gCDW−1 h−1) and d specific glucose uptake rate (mmol gCDW−1 h−1) at different dilution rates for S. cerevisiae ST938. Cultivations were carried out in triplicates at 30 °C and pH 5.5 under C-limited conditions. Virheet vastaavat kolmikertaisista viljelmistä johdettuja keskihajonnoja
S. cerevisiae ST938: n viljely C-rajoitusolosuhteissa osoitti siirtymistä eräajossa havaitusta pääasiassa fermentoivasta metaboliasta kemostaattien hengitystieaineenvaihduntaan, mikä näkyy suurempina biomassasatoina sekä vähäisenä etanolin ja glyserolin muodostumisena (Taulukko 1). Lisäksi pieniä määriä glukoosijäämää, joka on alle 0.Eri reaktoreiden effluxista otetuissa näytteissä havaittiin 1 mM, jolloin varmistettiin, että viljelmät eivät sisällä glukoosia. Vakaan tilan kemostaattiviljelmissä esiintyvät hiilirajoitteiset olosuhteet näyttivät edistävän 3 HP: n muodostumista, koska tämän tuotteen saanto oli 20-25 kertaa suurempi kuin eräviljelmässä määritetty saanto (Taulukko 1). Tämä johtuu mitä todennäköisimmin siitä, että hiili muuttuu tehokkaammin energiaksi ATP: n muodossa hengityksen aikana verrattuna käymiseen eräviljelmissä. On kiinnostavaa, että S. cerevisiae ST938 C-rajoitusolosuhteissa osoitti, että spesifisen kasvunopeuden hidastuessa voitiin havaita 3 hv: n hiilensaannin jatkuva kasvu, kun korkein 3 hv: n saanto oli 16, 6% c-mol laimennusnopeudella 0, 04 h-1 (kuva. 2 a). Biomassan 3 hv: n saanto on kymmenkertainen eräviljelmiin verrattuna, ja se nousi yli kaksinkertaiseksi 0,19−0,43 g gCDW−1: stä alentamalla laimennusnopeutta 0,21: stä 0,04 h-1: een (Taulukko 1). Suurin spesifinen tuottavuus 3-HP: lle oli noin 0,50 mmol gcdw−1 h-1 laimennusnopeuksilla 0,17 ja 0.21 h-1, joka on noin neljä kertaa suurempi kuin eräviljelmillä mitattu (Kuva. 2C, Taulukko 1). Spesifinen glukoosin kertymänopeus (qS) lisääntyi odotetusti, kun laimennusnopeus oli 0, 51 mmol: sta 2, 1 mmol: iin gcdw−1 h−1 (kuva. 2d). Nämä arvot ovat jopa 20 kertaa alhaisemmat kuin µmax: ssa mitattu spesifinen glukoosin kertymänopeus (10, 7 mmol gcdw−1 h−1) glukoosin rajoitetuissa kasvuolosuhteissa eräajotilassa (Taulukko 1). Substraatin ominaiskertymälle ja biomassan saannolle laimennusnopeudella 0 saadut arvot.09 h−1 (Taulukko 1) oli yhtäpitävä villin S. cerevisiae-tyypin kemostaattiviljelmistä saatujen tietojen kanssa laimennusnopeudella 0, 10 H−1, jolloin laimennusnopeus oli 1, 1 ja 1, 25 mmol gcdw−1 h−1 Sekä 0, 49 g g−1 . On havaittu, että biomassan saanto substraattia kohti kasvaa laimennusnopeuksien kasvaessa, mutta luonnonvaraisen S. cerevisiae-tyypin osalta tämä parametri on kuitenkin yleensä vakio alle Dcrit-arvon niin kauan kuin ylläpitoaineenvaihdunta muodostaa huomattavan hiilinielun, joka vähentää biomassan saantoa alhaisilla laimennusnopeuksilla . Jos kyseessä on S. cerevisiae-kanta ST938 näytti kuitenkin siltä, että hiili, joka ei ollut suunnattu biomassan muodostukseen, vaikutti jossain määrin sekä 3-HP: n että sivutuotteiden muodostumiseen.
johtopäätöksenä voidaan todeta, että jatkuvissa viljelmissä eri laimennusnopeuksilla koetut erot sekä S. cerevisiae-villityypin samankaltaisiin asetelmiin verrattuna viittaavat siihen, että integroidulla biosynteettisellä reitillä tuottaa 3-HP: ta on valtava vaikutus hiivan fysiologiaan, jota oletettavasti vahvistavat Adaptiiviset stressivasteet. Lopuksi C-rajoitettu kemostaatin viljely PIENIMMÄLLÄ laimennusnopeudella 0.04 h-1 sai aikaan suurimman 3 hv: n hiilituoton.
tutkittaessa typen ja fosfaatin rajoitusta 3 HP: n tuotoksen lisäämiseksi pienimuotoisissa kemostaattiviljelmissä
sen jälkeen, kun S. cerevisiae ST938 oli määritelty C-rajoitusolosuhteissa, tehtiin typen ja fosfaatin rajoituksia rajoittavia viljelyjä, koska ne ovat aiemmin osoittautuneet suotuisiksi erilaisten kotoperäisten ja ei-kotoperäisten metaboliittien tuotannolle eri eliöissä . Tässä tarkasteltiin vastaavia typen (N) ja fosforin (P) rajoituksen alaisia viljelmiä pienimuotoisessa jatkuvan viljelyn järjestelmässä (Kuva. 1) asetetulla laimennusnopeudella 0,04 h−1, joka oli aiemmin johtanut korkeimpaan 3-hevosvoiman hiilensaantiin C-rajoitetuissa olosuhteissa (Taulukko 1). St938 – hiivakannan n-ja P-rajoitetusta viljelmästä saadut parametrit ja vastaavat standardipoikkeamat on esitetty taulukossa 2.
N – ja P-rajoitetuissa viljelyolosuhteissa ei havaittu ammonium-tai fosfaattijäämiä eri reaktoreiden poistovedestä otetuissa näytteissä, jotta varmistettiin, että viljelmät ovat rajoitetut suhteessa vastaavaan substraattirajoitukseen (Taulukko 2). N – ja P-rajoituksessa havaittiin kuitenkin pieniä määriä 0,63 ja 0,19 mM: n glukoosijäämiä (Taulukko 2). C-rajoituksen aikana jäljellä oleva glukoosipitoisuus oli selvästi alle 0,1 mM (Taulukko 2). N – ja P-rajoituksia soveltavilla viljelmillä 3 hv: n saanto oli 17,7% C-mol ja 21,1% C-mol vastaavasti huomattavasti suurempi P-rajoitetuissa olosuhteissa verrattuna 3 hv: n hiilensaantiin 16,6% C-mol laskettuna C-rajoitetuissa viljelyolosuhteissa (taulukot 1 ja 2). Vastaavasti 3-HP: n spesifinen tuottavuus kasvoi merkittävästi N – ja P-rajoitetuilla viljelmillä, jotka paljastivat arvot 0,22 ja 0,23 mmol gcdw−1 h−1 verrattuna C−rajoitetuilla viljelmillä saavutettuun 0,17 mmol gcdw−1 h-1: een. Lisäksi 3 hv: n tuotot biomassasta olivat vertailukelpoiset arvot 0,43 ja 0.44 g gCDW – 1 C-ja N-rajoitusolosuhteissa, mutta P−rajoituksen tapauksessa havaittiin lisääntynyt saanto 0,50 g gCDW-1. Koska määrälliset sivutuotteiden kokonaispitoisuudet ovat pienet, ne ovat mitättömiä (Taulukko 2). Spesifisen substraatin kertymänopeuden (qS) arvot C – ja P-rajoitukselle olivat vertailukelpoisia, ja hieman kohonnut arvo 0, 61 mmol gcdw−1 h−1 saatiin N-rajoitetuissa olosuhteissa. Biomassasadot substraattia kohti n – ja p-rajoitetuissa olosuhteissa vastaavat arvot ovat 72,4 g mol−1 ja 75,2 g. Verrattuna biomassatuotokseen 78.4 g mol−1 saavutetaan C-rajoitetuissa olosuhteissa, nämä arvot ovat samanlaiset. Tutkimuksemme tukee sitä, että suuremmat tuotesadot substraattia kohti saavutettiin N – ja P-raja-olosuhteissa. Näin ollen vaikuttaa siltä, että korkeampi spesifinen substraatin kertymäaste N-rajoituksen tapauksessa yhdistettynä sivutuotteiden vähäiseen muodostumiseen suosi lopulta tuotteen muodostumista. P: tä rajoittavissa olosuhteissa muodostui vähiten sivutuotteita, jotka saattoivat suosia tuotesynteesiä. Voidaan edelleen spekuloida, että jotkin metaboliset muutokset mahdollisesti johtavat siihen, että hiilidioksidin vapautuminen hiilidioksidina vähenee.
yhteenvetona voidaan todeta, että kemostaattiviljelmillä N – ja P-rajoitusten mukaan 3 hv: n sadot ja erityiset tuotantonopeudet kasvoivat verrattuna C-rajoituksiin, ja P-rajoitus mahdollisti suurimmat tuotesadot.
pienimuotoisissa kemostaateissa mitattujen fysiologisten parametrien siirrettävyys 1-litraisissa bench-top bioreaktoreissa C-ja P-rajoitusolosuhteissa
koska tässä tutkimuksessa pyrittiin edelleen arvioimaan eri viljelyolosuhteissa mitattujen fysiologisten parametrien siirrettävyyttä ja vertailtavuutta, kemostaattiviljelmissä määritetyistä C – ja P-rajoitusolosuhteista saadut viljelyparametrit siirrettiin 1 – L sekoitettuihin penkki-latvareaktoreihin, jotka toimivat fed-batch-tilassa. Tämän lähestymistavan tarkoituksena oli pitää kemostaattiviljelmien ja eräviljelmien Keskeiset olosuhteet ja parametrit muuttumattomina vertailukelpoisuuden varmistamiseksi. Näissä olosuhteissa oli I) sama C:P-suhde kuin P: tä rajoittavissa kemostaattiviljelmissä, ii) identtiset prosessiolosuhteet, kuten pH ja lämpötila, ja iii) samat spesifiset kasvunopeudet kuin kemostaateissa, jotka saavutettiin eksponentiaalisella rehuprofiililla. Tuotteen titterin optimoimiseksi parametrit, joilla on maksimaalinen tuotteen saanto substraattia kohti kemostaattikokeista (D = 0.04 h – 1) alle C-ja P-rajoitus valittiin siirrettäväksi fed-eräjärjestelmään. Set-Upin teknisten rajoitusten vuoksi fed-eräviljelyn kasvunopeus oli 0,05 h−1, Mikä on hieman suurempi verrattuna kemostaattiviljelmien asetettuun laimennusnopeuteen 0,04 h-1. Fed-eräviljely koostui alkuerävaiheesta biomassan tuottamiseksi ja sen jälkeen eksponentiaalisesta, ravinnerajoitetusta ruokintavaiheesta kasvunopeuden säätelemiseksi. Ainoastaan ruokintavaihetta pidettiin merkityksellisenä jatkuvaviljelyjärjestelmästä saatujen parametrien siirrettävyyden arvioinnissa, koska ainoastaan tämän vaiheen aikana substraattipitoisuus kontrolloi ja siten rajoitti mikrobikasvustoa. S. cerevisiae ST938: n syötettyjen erien viljelyprofiilit, joissa sovelletaan C-ja P – rajoituksia, on esitetty kuvassa. 3, ja vastaavat viljelyparametrit ja niiden keskihajonta on esitetty taulukossa 3.
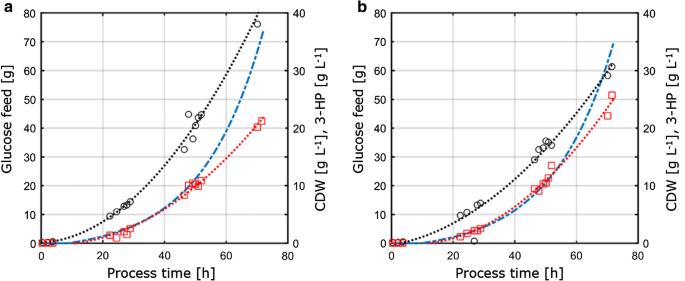
aerobinen fed-eräviljely S. cerevisiae ST938: n 1-L penkkifermenttereissä käyttäen eksponentiaalista ruokintaramppia d = 0, 05 h-1: ssä rajoittavan substraatin a hiilen tai b fosforin kanssa. Mustat ympyrät viittaavat biomassatitteriin, punaiset ruudut 3-HP: n titteriin ja sininen viiva reaktoreihin syötetyn glukoosin absoluuttiseen määrään
fed-eräviljelmissä biomassatuotos oli 77,9 g mol−1 C – ja p-rajoituksessa 67,7 g. C-rajoitettujen viljelmien biomassatuotos oli vertailukelpoinen kemostaatin ja fed-erien osalta, kun taas p-rajoitusten osalta biomassatuotos pieneni hieman Fed-erien käytön aikana (taulukot 2 ja 3), Mikä on molempien biomassatuotosten osalta Havaittujen keskihajonnojen sisällä. Hiilensaanti substraattia kohden 3-HP: lle oli 15,9% c-rajoituksessa, mikä on lähes identtinen verrattuna kemostaattijärjestelmässä laskettuun arvoon (Fig. 4 A). Tässä tutkimuksessa määritetyt 3-HP: n tuotot ovat edelleen lähellä sitä, että aiemmissa C-limited fed-erätutkimuksissa saatu hiilisaanto substraattia kohti pH 5, 0: ssa S. cerevisiae-kannalla, joka on suunniteltu käyttämään β-alaniinireittiä, ja 13%: ssa S. cerevisiae-kannalla, jossa käytetään malonyyli-CoA-reduktaasista riippuvaista reittiä . Escherichia coli-bakteerilla tehdyissä tutkimuksissa määritettiin korkeampi 3 HP: n hiilensaanti substraattia kohti β-alaniinireitin kautta, ja glukoosin arvo oli 42% eräruokinnassa tehdyissä viljelmissä .
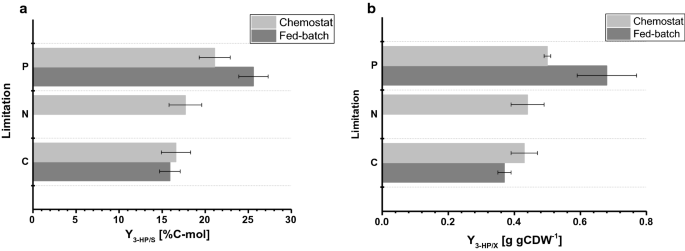
Viljelyparametrien Vertailu erilaisissa rajoituksissa, jotka on määritetty pienimuotoisissa kemostaateissa ja 1-litraisissa penkkipäällysreaktoreissa fed-eräajossa. 3 hv: n hiilensaanti (%C-mol) ja B 3 hv: n tuotto biomassasta (g gCDW-1). Virheet vastaavat kolmoisviljelmistä johdettuja keskihajonnoja
P-rajoitusolosuhteissa, mutta tutkimuksemme osoitti 3-HP: n hiilensaannin lisääntyneen huomattavasti 25,6% (Kuva. 4 A). Havaitut 3 hv: n tuotot biomassasta olivat 0,38 g gCDW−1 C – ja P-rajoitusolosuhteissa 0,65 g. C-rajoituksen osalta tämä vahvisti kemostaattijärjestelmässä havaitut arvot (Kuva. 4b). Koska 3-HP: n tuotto substraattia kohti kasvoi P-rajoitusolosuhteissa ja biomassan tuotto substraattia kohti laski hieman samaan aikaan, 3-HP: n tuotto biomassaa kohti kasvoi merkittävästi kemostaattikokeeseen verrattuna. Toisin kuin kemostaateissa suoritetuissa viljelyissä, etanolin tai glyserolin merkittävää kertymistä ei havaittu eräruokinnassa tehdyissä viljelmissä. C – ja P-raja-arvojen suljettu hiilitase osoitti, ettei muita (sivutuotteita)muodostu relevantteja määriä (Taulukko 3). Tämä sivutuotteiden spektrin ero saattaa selittää 3 HP: n tuotoksen lisääntymisen glukoosia kohti.
verrattuna kemostaateilla tehtyihin viljelmiin fed-eräviljelmissä määritetyt erityiset glukoosin kertymänopeudet osoittavat huomattavasti korkeammat arvot 0, 75 mmol gcdw−1 h−1 C – ja p-rajoitusolosuhteissa 0, 77 mmol hcdw-1 h-1. Kuten edellä mainittiin, syöttöprofiilin eksponentiaalisen kertoimen asetuspiste asetettiin 0,05 h−1: een ja vaihteli jopa 0,059 h-1: een, mikä johti 30-45% suurempaan spesifiseen kasvunopeuteen syötetyissä eräviljelmissä verrattuna kemostaattikokeisiin 0: lla.04 h−1 (Taulukko 3), mikä lisää spesifistä glukoosin kertymistä. C-rajoitetuissa fermentaatioissa glukoosin biomassatuotto oli kuitenkin vertailukelpoinen kemostaatin ja syötetyn erän viljelyn välillä. Vastaavasti 3 hv: n spesifiset tuotantonopeudet olivat koholla fed-batch-viljelmillä, vastaavasti 0,24 mmol gcdw−1 h−1 ja 0,38 mmol gcdw−1 h−1. 3 hv: n sato substraattia kohti oli vertailukelpoinen kemostaatin ja rehuerän viljelyn välillä, mikä viittaa siihen, että sato on kestävä viljelyparametri, joka kestää viljelyjärjestelyjen pienempien vaihtelujen aiheuttamia häiriöitä. Eri reaktoreista P-rajoitusolosuhteita varten otetuissa näytteissä ei havaittu fosfaatin eikä glukoosin jäämiä, mikä vahvisti viljelmien olevan rajoitettuja suhteessa vastaavaan rajoitukseen. Vastaavasti C-rajoituksen osalta näytteissä ei havaittu glukoosijäämiä.
tässä esitettiin parannuksia pienimuotoiseen kemostaattijärjestelmään, jonka aiemmin kehittivät Klein et al. , käsitti muun muassa reaktoreiden määrän lisäämisen 24: ään ja viljelytilavuuden vähentämisen 6,5 millilitraan. Tämä lisääntynyt paralleliaatioaste tekee järjestelmästä sopivan suurikapasiteettisen seulontatyökalun, koska useat vastaavat pienimuotoiset kemostaattijärjestelmät toimivat suuremmilla työskentelytilavuuksilla ja pienemmällä reaktorimäärällä: cosbios (continuous parallel shaken bioreactor)-järjestelmässä käytetään kuutta kahdeksaa rinnakkaista viljelyastiaa, joiden kantotilavuus on 20-25 mL . For continuous cultivations modified single-use stuffed-tank bioreactor system (bioREACTOR, 2mag AG) toimii kahdeksalla rinnakkaisella Reaktorilla 10 mL: n työskentelytilavuudella . Bergenholmin ym.kehittämä mini-chemostat (MC) – järjestelmä. koostuu 16 rinnakkaisesta reaktorista ja vaatii 40 mL: n työskentelytilavuuden. Järjestelmämme soveltuu siis hyvin mikrobien suorituskyvyn yksinkertaiseen ja kustannustehokkaaseen seulontaan jatkuvassa tilassa. Tällä järjestelmällä voisi kuitenkin olla merkitystä, jos se tarkkailisi lisäparametreja, kuten pakokaasua, jotta hiilijakauma voitaisiin analysoida yksityiskohtaisesti, mikä on erityisen tärkeää fysiologisten kantojen luonnehdinnan kannalta.
koska tutkimuksemme ehdotti N – ja P-rajoituksen olevan suotuisa 3-HP: n muodostumiselle S. cerevisiaessa, tuleviin kokeisiin voisi sisältyä kasvunopeuksien lisäseulonta N – ja P-rajoituksella, jotta löydettäisiin optimaalinen 3-HP: n tuotanto kullakin rajoituksella. Tutkimuksemme osoitti edelleen fysiologisten parametrien siirrettävyyden kemostaatista eräviljelyihin. Tätä pidetään mahdollisena vain, jos myrkyllisiä tai inhiboivia yhdisteitä ei kerry syötetyn erän viljelyn aikana, koska fysiologiset parametrit muuttuisivat ajan myötä ja poikkeaisivat kemostaattien hankituista parametreista. Yleisenä lähestymistapana kemostaattikokeet voivat toimia työkaluna mahdollisen myrkyllisen tai inhiboivan yhdisteen vaikutuksen tutkimiseen lisäämällä ainetta itse rehuun. Koska kaikki muut parametrit ovat vakio, yhdisteen ja sen konsentraation vaikutusta voidaan suoraan arvioida ja arvioida. Tutkimuksessamme ei havaittu sivutuotteenmuodostusta syötetyn erän viljelmillä ja oletettavasti samanlainen stressireaktio aiheutui heikkojen happojen vuoksi, mikä mahdollisti fysiologisten olosuhteiden pienimuotoisen kemostaatin samankaltaisuuden kuin syötetyn erän olosuhteissa.
yhteenvetona tämä tutkimus osoitti, että C – ja P-raja-olosuhteiden käsitettä 3-HP: n tuotannossa tutkittiin uudessa parallelized chemostat-viljelyjärjestelmässä, ja se voitiin onnistuneesti siirtää 1-L-penkki-top-bioreaktoreihin, jotka toimivat fed-eräajotilassa. Sen vuoksi kemostaateissa hankittuja fysiologisia parametreja voidaan käyttää syöttöerien viljelyjen suunnittelussa ja suorituskyvyn arvioinnissa käyttämällä prosessin asetukseen perustuvia parametreja.