
glykolyysi
glykolyysin alkupuoli käyttää glukoosin fosforylaatiossa kahta ATP-molekyyliä, jotka sitten jakautuvat kahdeksi kolmihiiliseksi molekyyliksi.
glykolyysin jälkipuoliskolla fosforylaatio tapahtuu ilman ATP-sijoitusta (Vaihe 6) ja muodostuu kaksi NADH-ja neljä ATP-molekyyliä glukoosia kohti.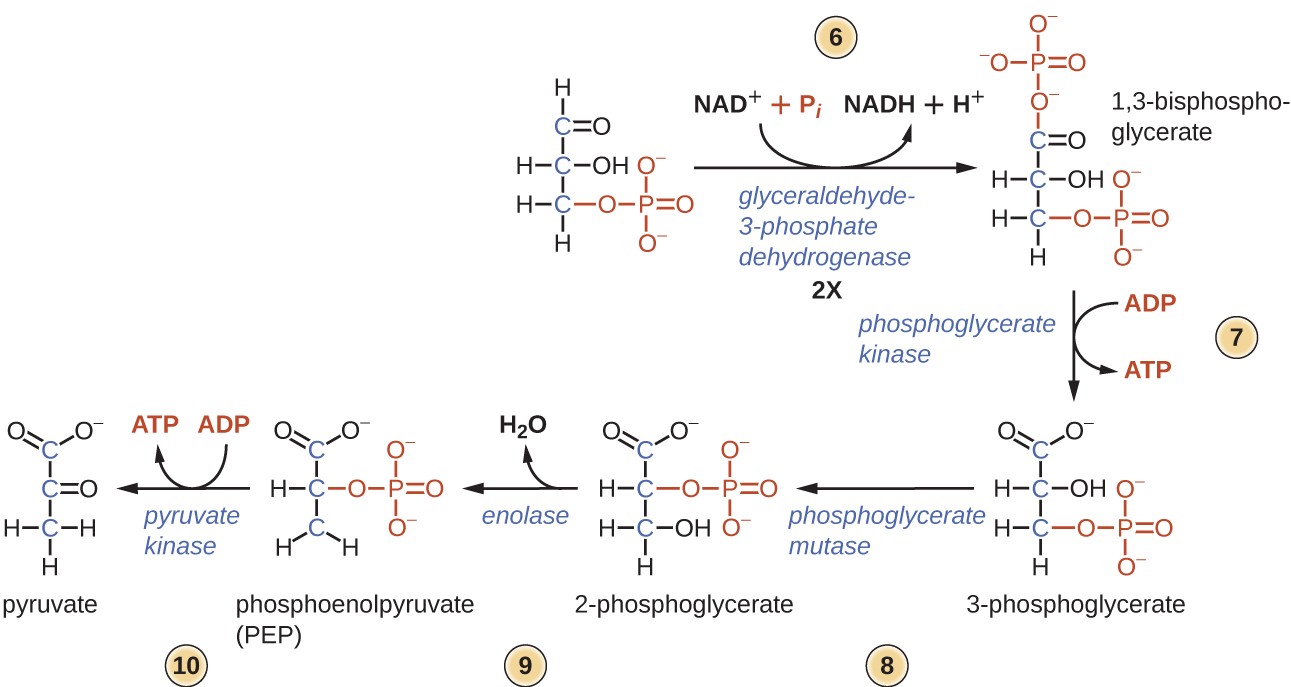
Entner–Doudoroff–reitti
Entner-Doudoroff-reitti on metaboliareitti, joka muuttaa glukoosin etanoliksi ja yhdistää yhden ATP: n.
Pentoosifosfaattireitti
pentoosifosfaattireitti, jota kutsutaan myös fosfoglukonaattireitiksi ja heksoosimonofosfaattisunttireitiksi, on glykolyysin suuntainen metaboliareitti, joka tuottaa NADPH: ta ja viiden hiilen sokereita sekä riboosi-5-fosfaattia, joka on glukoosin nukleotidien synteesin esiaste.
TCA Cycle
In this transition reaction, a multi-enzyme complex converts pyruvate into one acetyl (2C) group plus one carbon dioxide (CO2). Asetyyliryhmä kiinnittyy koentsyymi-A: n kantajaan, joka kuljettaa asetyyliryhmän Krebs-syklin paikalle. Prosessissa muodostuu yksi NADH-molekyyli.
sitruunahappokierrossa asetyyli-CoA: n asetyyliryhmä kiinnittyy nelihiiliseen oksaloasetaattimolekyyliin muodostaen kuusihiilisen sitraattimolekyylin. Askelsarjan kautta sitraatti hapetetaan, jolloin jokaisesta sykliin syötetystä asetyyliryhmästä vapautuu kaksi hiilidioksidimolekyyliä. Prosessissa substraattitason fosforylaatiolla tuotetaan kolme NADH: ta, yksi FADH2: ta ja yksi ATP: tä tai GTP: tä (solutyypistä riippuen). Koska sitruunahappokierron lopputuote on myös ensimmäinen reaktantti, sykli jatkuu jatkuvasti, kun reaktantteja on riittävästi. (luotto: ”Yikrazuul”/Wikimedia Commons)
Beetahapetus
rasvahappojen hapettumisen aikana triglyseridit voidaan jakaa 2C-asetyyliryhmiin, jotka pääsevät Krebs-sykliin ja joita voidaan käyttää energianlähteenä glukoositasojen ollessa alhaiset.
oksidatiivinen fosforylaatio
elektroninsiirtoketju on sarja elektroninkantajia ja ionipumppuja, joita käytetään H+ – ionien pumppaamiseen kalvon poikki. H+ virtaa sitten takaisin kalvon läpi ATP-syntaasin avulla, joka katalysoi ATP: n muodostumista. Elektroninsiirtoketjun sijainti on eukaryoottisolujen sisempi mitokondriomatriisi ja prokaryoottisolujen sytoplasmakalvosto.
Calvin-Bensonin sykli
Calvin-Bensonin syklissä on kolme vaihetta. Vaiheessa 1 rubisco-entsyymi yhdistää hiilidioksidin orgaaniseksi molekyyliksi, 3-PGA: ksi. Vaiheessa 2 orgaaninen molekyyli pelkistetään NADPH: n toimittamien elektronien avulla. Vaiheessa 3 kierron aloittava molekyyli RuBP regeneroidaan niin, että sykli voi jatkua. Vain yksi hiilidioksidimolekyyli on sisällytetty kerrallaan, joten sykli on suoritettava kolme kertaa yhden kolmihiilisen GA3P-molekyylin tuottamiseksi ja kuusi kertaa kuusihiilisen glukoosimolekyylin tuottamiseksi.