
solun Adheesioproteiinit
solun ja solun välinen adheesioproteiini on selektiivinen prosessi, jossa solut kiinnittyvät vain tiettyihin muihin soluihin. Tämä selektiivisyys osoitettiin ensimmäisen kerran klassisissa alkionkehitystutkimuksissa, jotka osoittivat, että solut yhdestä kudoksesta (esim.maksa) kiinnittyvät nimenomaan saman kudoksen soluihin eikä eri kudoksen soluihin (esim. aivot). Tällaisen selektiivisen solun ja solun välisen adheesion välittäjinä toimivat transmembraaniproteiinit, joita kutsutaan solun adheesio-molekyyleiksi, jotka voidaan jakaa neljään pääryhmään: selektiineihin, integriineihin, immunoglobuliinin (IG) superperheeseen (jotka on nimetty siten, koska ne sisältävät immunoglobuliineja muistuttavia rakenteellisia domeeneja) ja kadheriineihin (taulukko 12.3). Selektiinien, integriinien ja kadheriinien välittämä solujen adheesiota vaatii Ca2+ tai Mg2+, joten monet solujen väliset liima – vuorovaikutukset ovat Ca2+-tai Mg2+ – riippuvaisia.
taulukko 12.3
solun Adheesiomolekyylit.
selektiinit välittävät leukosyyttien ja endoteelisolujen tai verihiutaleiden välisiä ohimeneviä yhteisvaikutuksia. Selektiiniperheessä on kolme jäsentä: l-selektiini, joka ilmaistaan valkosoluilla; e-selektiini, joka ilmaistaan endoteelisoluilla; ja p-selektiini, joka ilmaistaan verihiutaleilla. Kuten aiemmin tässä luvussa on käsitelty, selektiinit tunnistavat solupinnan hiilihydraatit (KS.Kuva 12.14). Yksi niiden kriittisistä rooleista on aloittaa leukosyyttien ja endoteelisolujen väliset vuorovaikutukset leukosyyttien siirtymisen aikana verenkierrosta kudostulehduspaikoille (Kuva 12.62). Selektiinit välittävät leukosyyttien alkuperäistä adheesiota endoteelisoluihin. Tätä seuraa vakaampien adheesioiden muodostuminen, jossa leukosyyttien pinnalla olevat integriinit sitoutuvat solujen välisiin adheesiomolekyyleihin (ICAMs), jotka ovat endoteelisolujen pinnalla ilmentyvän IG-superperheen jäseniä. Lujasti kiinnittyneet leukosyytit pystyvät sitten tunkeutumaan kapillaarien seinämiin ja siirtymään taustalla olevaan kudokseen endoteelisolujen välillä.

Kuva 12, 62
leukosyyttien ja endoteelisolujen välinen adheesio. Leukosyytit poistuvat verenkierrosta kudostulehduksen kohdissa vuorovaikutuksessa kapillaariseinämien endoteelisolujen kanssa. Tämän yhteisvaikutuksen ensimmäinen vaihe on leukosyyttien selektiinien (more…)
ikaamien sitoutuminen integriineihin on esimerkki heterofiilisestä vuorovaikutuksesta, jossa yhden solun pinnalla oleva adheesiomolekyyli (esim.ikaami) tunnistaa toisen solun pinnalla olevan eri molekyylin (esim. integriini). Muut IG-superperheen jäsenet välittävät homofiilisia vuorovaikutuksia, joissa yhden solun pinnalla oleva adheesiomolekyyli sitoutuu samaan molekyyliin toisen solun pinnalla. Tällainen homofiilinen sitoutuminen johtaa selektiiviseen adheesioon samantyyppisten solujen välillä. Esimerkiksi hermosolujen adheesiomolekyylit (N-kamerat) ovat hermosoluissa ilmenevän IG-superperheen jäseniä, ja N-kameroiden välinen homofiilinen sitoutuminen edistää hermosolujen välisten selektiivisten assosiaatioiden muodostumista kehityksen aikana. Ig-superperheessä on yli 100 jäsentä, jotka välittävät erilaisia solujen ja solujen välisiä vuorovaikutuksia.
solujen neljännessä adheesiomolekyyliryhmässä, kadheriineissa, esiintyy myös homofiilisia sitoutumisominaisuuksia. Ne eivät osallistu ainoastaan alkiosolujen väliseen valikoivaan adheesioon, vaan ne ovat myös pääasiallisesti vastuussa vakaiden liitosten muodostumisesta solujen välille kudoksissa. Esimerkiksi E-kadheriini ilmaistaan epiteelisoluilla, joten E-cadheriinien väliset homofiiliset vuorovaikutukset johtavat epiteelisolujen selektiiviseen kiinnittymiseen toisiinsa. On huomionarvoista, että E-cadherinin menetys voi johtaa epiteelisoluista johtuvien syöpien kehittymiseen, mikä havainnollistaa solujen ja solujen vuorovaikutusten merkitystä solujen käyttäytymisen säätelyssä. Cadherinin perheen eri jäsenet, kuten n-cadheriini (neural cadherini) ja P-cadheriini (istukkakadheriini), välittävät muiden solutyyppien selektiivistä adheesiota.
on tunnistettu parikymmentä erilaista klassista cadherinia, kuten E-cadherinia. Lisäksi erillinen cadheriinien alaheimo, protokadheriinit, ilmenee keskushermostossa, jossa niillä näyttää olevan rooli hermosolujen välisessä adheesiossa synapseissa. Kiehtovasti eri neuronit näyttävät ilmaisevan erilaisia protokadheriineja, mikä viittaa siihen, että protokadheriineilla voi olla rooli erityisten yhteyksien luomisessa neuronien välille. Noin 50 ihmisen protokadheriinigeeniä on tunnistettu ja osoitettu järjestäytyneen kolmeen geeniklusteriin. Jokainen klusteri sisältää useita eksoneja, jotka koodaavat n-terminaalisia solunulkoisia ja transmembraanisia protokadheriinidomeeneja, mutta vain yhden kolmen eksonin joukon, jotka koodaavat C-terminaalista sytoplasmodomeenia (Kuva 12.63). Protokadheriinigeeniklusterit näyttävät siis koostuvan muuttuvasta alueesta, joka koodaa useita solunulkoisia ja transmembraanisia domeeneja, jotka ovat yhteydessä yhtä sytoplasmadomeenia koodaavaan vakioalueeseen. Tämä protokadheriinigeenien organisaatio muistuttaa silmiinpistävästi immunoglobuliini-ja t-solureseptorigeenien organisaatiota (KS.kuvat 5.42 ja 5.43), jossa useita muuttuvia alueeksoneja yhdistyy yhdeksi vakioalueeksoniksi. Immunoglobuliini-ja T-solureseptorigeeneissä tämä tapahtuu DNA: n uudelleenjärjestelyjen seurauksena, jotka synnyttävät monimuotoisuutta immuunijärjestelmässä. On vielä selvitettävä, liittyvätkö protokadheriinien vaihtelevat ja pysyvät alueet toisiinsa DNA-tai RNA-tasolla (esimerkiksi vaihtoehtoisella yhteenliittämisellä) ja missä määrin protokadheriinigeenien uudelleenjärjestelyt voivat edistää erityisten synaptisten yhteyksien syntymistä aivoissa.
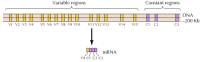
Kuva 12.63
protokadheriinigeeniklusterien järjestäminen. Ihmisen protokadheriini-geenit ovat järjestäytyneet kolmeen ryppääseen. Havainnollistetussa klusterissa 15 eri muuttuvaa aluetta, jotka koodaavat solunulkoisia ja transmembraanisia domeeneja, liittyvät yhteen vakioalueeseen, (lisää…)
toisin kuin edellisessä jaksossa käsitellyt vakaat solumatriisiliitokset, selektiinien, integriinien ja Ig-superperheen jäsenten välittämät solujen ja solujen väliset interaktiot ovat ohimeneviä kiinnikkeitä, joissa viereisten solujen tukirangat eivät ole yhteydessä toisiinsa. Vakaat adheesioliitokset, joihin liittyy viereisten solujen sytoskeletonit, välittyvät sen sijaan cadheriinien välityksellä. Kuten luvussa 11 käsitellään, näitä solun ja solun välisiä liitoksia on kahdenlaisia: adhereeniliitoksia ja desmosomeja, joissa kadheriinit tai niihin liittyvät proteiinit (desmogleiinit ja desmokolliinit) liittyvät aktiinikimppuihin ja välifilamentteihin (Kuva 12.64). Kadheriinien rooli viereisten solujen sytoskeletonien yhdistämisessä on siten analoginen integriinien kanssa vakaiden liitosten muodostamisessa solujen ja solunulkoisen matriisin välille.

Kuva 12.64
cadheriinien välittämät stabiilit soluliitokset. Kadheriinien väliset homofiiliset vuorovaikutukset välittävät kahdenlaisia stabiileja soluliitoksia. Adhereenien liitoksissa kadheriinit liittyvät aktiinifilamenttien nippuihin kateniinien kautta(ks…)