Sistema mejorado de cultivo continuo a pequeña escala
En este estudio, mostramos nuevos avances y posibilidades de aplicación de un sistema de cultivo continuo a pequeña escala, desarrollado previamente por Klein et al. , para un mayor grado de paralelización y un mejor manejo, así como la supervisión de los reactores individuales. Los aspectos clave de los cambios realizados en el sistema incluyen un aumento del conjunto de recipientes de cultivo paralelos de 8 a 24 reactores y una disminución del volumen de trabajo de 10 a 6,5 ml. El sistema actual consiste además en tapas hechas a medida que albergan cuatro puertos fijos utilizados para la aireación, el suministro de medios, la extracción de caldo, así como la inoculación o el muestreo (Fig. 1). Además de los cuatro puertos, se inserta una sonda de DO en forma de varilla óptica a través de la tapa para monitorear el DO sin alterar el proceso de cultivo, y de esta manera reemplazar el punto del sensor fluorescente de oxígeno de la configuración anterior . El baño de agua, que en la configuración anterior mantenía una temperatura de cultivo constante, fue reemplazado por un bloque calefactor de aluminio hecho a medida, que se fusiona con una unidad agitadora de microplacas. Como la versión anterior del sistema de biorreactores a pequeña escala se validó utilizando la levadura de fisión Schizosaccharomyces pombe, presentamos aquí la configuración de cultivo mejorada para cultivos de S. cerevisiae.
Se realizaron los pasos operativos básicos, así como los ajustes de las tasas de dilución mediante la selección del diámetro del tubo y la tasa de bomba apropiados de la bomba de flujo de medios, como se describió anteriormente . Aquí, el peso del contenido líquido de cada biorreactor se determinó gravimétricamente al final del cultivo, lo que permitió el cálculo preciso de la tasa de dilución respectiva con un 5.desviación del 1%. El caldo de cultivo y la fase de gas del reactor se eliminaron a través del mismo puerto de la tapa del reactor utilizando la bomba de eflujo (Fig. 1). Para todos los experimentos de cultivo se utilizaron caudales de bombeo de salida de 7,5 mL min−1. La velocidad de la bomba de salida era muy superior a la velocidad de la bomba de alimentación, generando una ligera presión negativa dentro del recipiente de cultivo. Esta diferencia de presión dio lugar a la entrada de aire a través del puerto de aireación. El coeficiente de transferencia de masa de oxígeno kLa promedio alcanzado fue de 110 h-1, lo que permitió niveles de DO muy superiores al 30% de saturación durante todo el proceso de cultivo. El pH no fue monitoreado en línea ni controlado durante el cultivo, ya que el medio se ajustó a priori a un pH de 6,0, lo que resultó en un pH final de 5,5 en el caldo de cultivo. El pH se midió en línea diariamente a partir del flujo de salida de los reactores y después de la cosecha. El pH se mantuvo constante tan pronto como se alcanzó el estado estacionario y el efluente del reactor mostró una desviación menor de 0,1 unidades de pH (datos no mostrados).
Explorando la producción de 3 HP en quimiostatos a pequeña escala a diferentes velocidades de dilución en condiciones limitantes de C
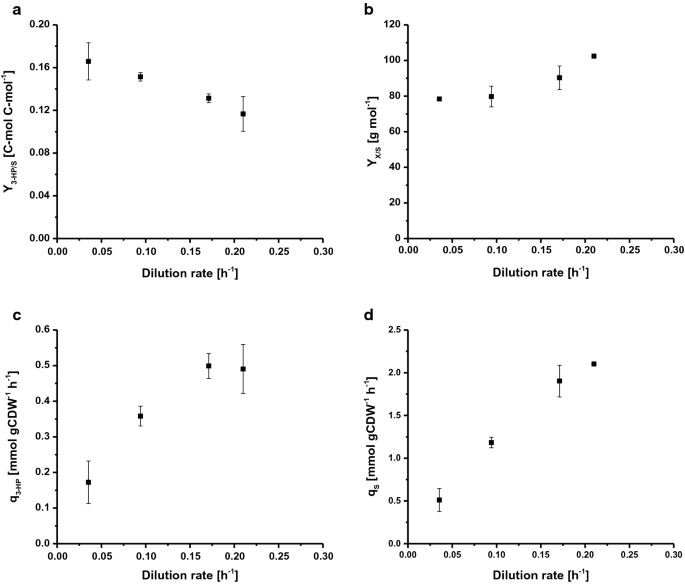
Para determinar la tasa de crecimiento específico máximo (µmáx), S. cerevisiae ST938 se cultivó en lotes aplicando disponibilidad excesiva de nutrientes. Utilizando glucosa como fuente de carbono, el µmáx fue de 0,265 h−1, el rendimiento de biomasa fue de 24,9 g mol-1 y el rendimiento de carbono de 3 HP en glucosa fue de 0,6% C-mol (Tabla 1). En este modo de cultivo, la mayor parte del carbono se metabolizan el etanol y CO2 , como glicolítica alta flujos de tipo salvaje S. las cerevisiae están fuertemente vinculadas a la fermentación alcohólica . Los cultivos de quimiostatos se controlan mediante el suministro de un sustrato que limita el crecimiento. En estado estacionario, S. cerevisiae de tipo silvestre no produce cantidades significativas de metabolitos de desbordamiento por debajo de una determinada tasa de crecimiento, marcando la tasa de dilución crítica (Dcrit (h−1)) debido a la falta de acumulación de fuente de carbono. En o por encima de esta tasa de dilución crítica, la fuente de carbono se acumula en el reactor y se activa el metabolismo de desbordamiento, lo que resulta en la producción de varios subproductos, como etanol, acetato y pequeñas cantidades de ácidos orgánicos . En consecuencia, se utilizaron cultivos continuos para investigar la relación entre la tasa de crecimiento y la formación de productos de 3 HP específicos de biomasa en S. cerevisiae ST938. Las cuatro tasas de dilución diferentes D (h−1): 0,04, 0,09, 0,17 y 0,21 se eligieron por debajo del µmáx de la cepa ST938 de S. cerevisiae determinada en el cultivo por lotes. Se investigó el rendimiento de biomasa en glucosa, así como el rendimiento de 3 HP, la tasa de producción específica de 3 HP y la tasa de absorción de sustrato específico (Fig. 2a a d, cuadro 1). En el cuadro 1 se resumen todos los parámetros de cultivo pertinentes, es decir, rendimientos y tasas con las desviaciones estándar respectivas.
Tabla 1 parámetros de Cultivo de S. cerevisiae ST938 crecido en culturas continuas en C-condiciones limitadas
Fig. 2
Selected rates and yields for C-limited aerobic chemostat cultivations of S. cerevisiae ST938 a 3-HP yield (C-mol C-mol−1) on glucose, b biomass yield on glucose (g mol−1), c specific 3-HP production rate (mmol gCDW−1 h−1) and d specific glucose uptake rate (mmol gCDW−1 h−1) at different dilution rates for S. cerevisiae ST938. Cultivations were carried out in triplicates at 30 °C and pH 5.5 under C-limited conditions. Los errores corresponden a desviaciones estándar derivadas de cultivos triplicados
El cultivo de S. cerevisiae ST938 en condiciones limitantes de C mostró un cambio del metabolismo predominantemente fermentativo observado en modo por lotes a un metabolismo respiratorio en quimiostáticos, que se refleja en mayores rendimientos de biomasa, así como en una formación insignificante de etanol y glicerol (Tabla 1). Además, cantidades menores de glucosa residual por debajo de 0.Se detectaron 1 mm en las muestras tomadas del flujo de salida de los diferentes reactores, verificando que los cultivos estaban limitados a glucosa. Las condiciones limitadas en carbono encontradas en los cultivos de quimiostatos en estado estacionario parecían promover la formación de 3 HP, ya que los rendimientos de este producto eran de 20 a 25 veces superiores al rendimiento determinado en el cultivo por lotes (Tabla 1). Esto es probablemente debido a la conversión más eficiente de carbono en energía en forma de ATP durante la respiración en comparación con la fermentación en cultivos por lotes. Curiosamente, los cultivos de S. cerevisiae ST938 en condiciones limitantes de C reveló que con tasas de crecimiento específicas decrecientes se podía observar un aumento constante en los rendimientos de carbono de 3 HP, con el rendimiento más alto de 3 HP de 16,6% C-mol observado a una tasa de dilución de 0,04 h-1 (Fig. 2a). El rendimiento de 3 HP en biomasa es diez veces mayor que el rendimiento en cultivos por lotes y aumentó más del doble de 0,19 a 0,43 g de gCDW-1 al reducir la tasa de dilución de 0,21 a 0,04 h−1 (Tabla 1). La productividad específica máxima para 3 HP de aproximadamente 0,50 mmol de gCDW-1 h−1 se observó a tasas de dilución de 0,17 y 0.21 h-1, que es aproximadamente cuatro veces mayor que la medida en cultivos por lotes (Fig. 2c, cuadro 1). Como era de esperar, la tasa específica de absorción de glucosa (qS) aumentó con tasas de dilución más altas de 0,51 a 2,1 mmol gCDW−1 h−1 (Fig. 2d). Estos valores están hasta 20 veces por debajo de la tasa de absorción de glucosa específica máxima de 10,7 mmol gCDW−1 h−1 observada en µmáx en condiciones de crecimiento limitado a la glucosa en modo por lotes (Tabla 1). Valores obtenidos para la tasa de absorción del sustrato específico y el rendimiento de biomasa a una tasa de dilución de 0.09 h-1 (Tabla 1) coincidieron con los datos de cultivos de quimiostatos con S. cerevisiae de tipo silvestre a una velocidad de dilución de 0,10 h−1, indicando 1,1 y 1,25 mmol gCDW−1 h−1, así como 0,49 g g−1, respectivamente . Se ha encontrado que el rendimiento de biomasa por sustrato aumenta con el aumento de las tasas de dilución, para S. cerevisiae de tipo silvestre, sin embargo, este parámetro generalmente es constante por debajo de Dcrit siempre que el metabolismo de mantenimiento constituya un sumidero de carbono considerable que reduzca el rendimiento de biomasa a bajas tasas de dilución . En el caso de S. cerevisiae cepa ST938, sin embargo, parecía que el carbono, que no estaba dirigido a la formación de biomasa, estaba contribuyendo en cierta medida a la formación de 3 HP y subproductos.
Para concluir, las diferencias experimentadas en cultivos continuos a diferentes tasas de dilución, así como en comparación con configuraciones similares con S. cerevisiae de tipo silvestre, sugieren que la vía biosintética integrada para producir 3-HP tiene un tremendo impacto en la fisiología de la levadura, que presumiblemente se amplifica por respuestas de estrés adaptativas. Finalmente, C-limitó el cultivo de quimiostatos a la tasa de dilución más baja de 0.04 h-1 resultó en el mayor rendimiento de carbono de 3 HP.
Tras caracterizar S. cerevisiae ST938 bajo condiciones limitantes de C, se realizaron cultivos bajo condiciones limitantes de nitrógeno y fosfato, ya que se ha demostrado previamente que son favorables para la producción de varios metabolitos nativos y no nativos en diferentes organismos . Aquí, se investigaron los cultivos correspondientes bajo limitación de nitrógeno (N) y fósforo (P) en el sistema de cultivo continuo a pequeña escala (Fig. 1) con una tasa de dilución establecida de 0,04 h−1, que anteriormente había dado como resultado el mayor rendimiento de carbono de 3 HP en condiciones C limitadas (Tabla 1). Los parámetros y las respectivas desviaciones estándar de los cultivos N y P limitados de la cepa de levadura ST938 se resumen en la Tabla 2.
Tabla 2 Parámetros de cultivo de S. cerevisiae ST938 cultivados en cultivos continuos a una velocidad de dilución de 0.04 h – 1 en condiciones limitadas de carbono (C), nitrógeno (N) y fósforo (P)
Para condiciones de cultivo limitadas de N y P, no se detectaron residuos de amonio o fosfato en las muestras tomadas del flujo de salida de los diferentes reactores, verificando que los cultivos se limitarán con respecto a la limitación de sustrato correspondiente (Tabla 2). Sin embargo, se detectaron cantidades menores de glucosa residual en el rango de 0,63 y 0,19 mm bajo limitación de N y P (Tabla 2). Durante la limitación de C, los niveles residuales de glucosa estuvieron muy por debajo de 0,1 mm (Tabla 2). Los cultivos que aplicaban limitaciones de N y P revelaron un rendimiento de 3 HP de 17,7 y 21,1% C-mol, respectivamente, mostrando un valor significativamente mayor bajo condiciones de P-limitado en comparación con el rendimiento de carbono de 3 HP de 16,6% C-mol calculado para condiciones de cultivo C-limitado (Tablas 1, 2). De manera similar, la productividad específica para 3 HP se incrementó significativamente para cultivos N y P limitados, revelando valores de 0,22 y 0,23 mmol gCDW-1 h – 1 en comparación con 0,17 mmol gCDW-1 h−1 alcanzados en cultivos C limitados. Además, los rendimientos de 3 HP en biomasa mostraron valores comparables de 0,43 y 0.44 g de gCDW – 1 para condiciones limitantes de C y N, sin embargo, en caso de limitación de P, se observó un aumento del rendimiento de 0,50 g de gCDW-1. Debido a las bajas concentraciones globales de subproductos cuantificados, éstas son insignificantes (cuadro 2). Los valores de la tasa de absorción de sustrato específico (qS) para la limitación de C y P fueron comparables y se obtuvo un valor ligeramente elevado de 0,61 mmol gCDW – 1 h-1 en condiciones N limitadas. Rendimientos de biomasa por sustrato revelados para condiciones limitadas de N y P valores similares de 72,4 y 75,2 g mol-1, respectivamente. Comparado con el rendimiento de biomasa de 78.4 g de mol−1 alcanzados en condiciones C-limitadas, estos valores son similares. Nuestro estudio apoya la justificación de que se lograron mayores rendimientos de producto por sustrato en condiciones limitantes de N y P. En consecuencia, parece que una mayor tasa de absorción de sustrato específico en caso de limitación de N combinada con la formación mínima de subproductos en última instancia favoreció la formación del producto. Bajo condiciones limitantes de P se formaron las menores cantidades de subproductos, lo que podría favorecer la síntesis de productos. Además, se puede especular que algunos cambios metabólicos posiblemente conduzcan a una menor cantidad de carbono que se libera como CO2.
En resumen, los cultivos de quimiostatos bajo limitación de N y P revelaron un aumento de los rendimientos de 3 HP y tasas de producción específicas en comparación con las condiciones limitadas por C, con la limitación de P permitiendo los rendimientos más altos del producto.
Transferibilidad de los parámetros fisiológicos medidos en quimiostatos de pequeña escala a cultivos alimentados por lotes en biorreactores de sobremesa de 1 L en condiciones limitantes de C y P
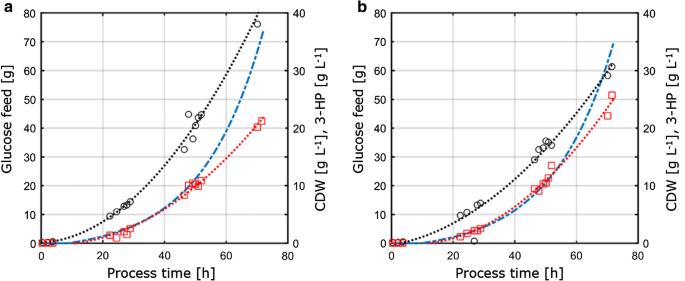
Dado que este estudio tuvo como objetivo evaluar la transferibilidad y comparabilidad de los parámetros fisiológicos medidos en diferentes condiciones de cultivo, los parámetros de cultivo obtenidos de las condiciones limitantes de C y P determinadas en cultivos de quimiostatos se transfirieron a reactores de sobremesa con agitación de 1 L que funcionan en modo alimentado por lotes. El concepto de este enfoque era, por lo tanto, mantener constantes las condiciones y los parámetros clave para los cultivos de quimiostatos y de lotes alimentados a fin de garantizar la comparabilidad. Estas condiciones comprendían (i) la misma relación C:P que se aplica en los cultivos de quimiostatos limitantes de P, (ii) las mismas condiciones de proceso, como el pH y la temperatura, y (iii) las mismas tasas de crecimiento específicas que se aplican en los quimiostatos, logradas a través de un perfil de alimentación exponencial. Para optimizar el título del producto, los parámetros con el rendimiento máximo del producto por sustrato de los experimentos de quimiostatos (D = 0.04 h-1) bajo limitación C – y P-fueron elegidos para la transferencia al sistema de alimentación por lotes. Debido a restricciones técnicas en la configuración, los cultivos por lotes alimentados se llevaron a cabo a una tasa de crecimiento de 0,05 h-1, que es ligeramente mayor en comparación con la tasa de dilución establecida de 0,04 h-1 en cultivos de quimiostatos. El cultivo por lotes con alimento consistió en una fase inicial de lotes para generar biomasa, seguida de una fase de alimentación exponencial con nutrientes limitados para controlar la tasa de crecimiento. Solo la fase de alimentación se consideró relevante para la evaluación de la transferibilidad de los parámetros obtenidos del sistema de cultivo continuo, ya que únicamente durante esta fase la concentración del sustrato controlaba y limitaba el crecimiento microbiano. Los perfiles de cultivo por lotes de S. cerevisiae ST938 aplicando condiciones limitantes de C y P se muestran en la Fig. 3, y los parámetros de cultivo correspondientes con sus respectivas desviaciones estándar se resumen en la Tabla 3.
Fig. 3
Tabla 3 Parámetros de cultivo de S. cerevisiae ST938 cultivado en modo de lote alimentado en biorreactores de sobremesa de 1 L en condiciones limitantes de carbono (C) y fósforo (P)
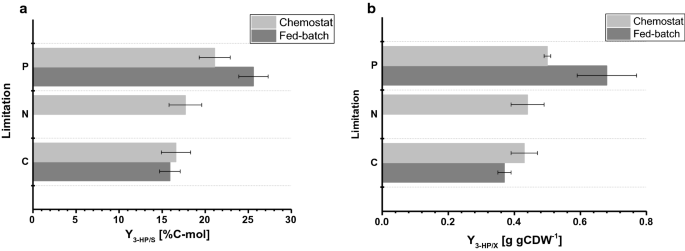
El rendimiento de biomasa observado en los cultivos de lote alimentado fue de 77,9 y 67,7 g mol-1 para limitación de C y P, respectivamente. El rendimiento de biomasa para cultivos limitados por C fue comparable entre el quimiostato y la operación por lotes alimentados, mientras que para la limitación por P, se observó una ligera disminución del rendimiento de biomasa durante la operación por lotes alimentados (Tablas 2, 3), que está dentro de las desviaciones estándar observadas para ambos rendimientos de biomasa. El rendimiento de carbono por sustrato para 3 HP fue de 15,9% para la limitación de C, que es casi idéntico en comparación con el valor calculado en el sistema de quimiostatos (Fig. 4a). Los rendimientos de 3 HP determinados en este estudio están además en estrecha concordancia con un rendimiento de carbono por sustrato del 14% obtenido en estudios previos de lotes alimentados con C limitada a pH 5,0 con una cepa de S. cerevisiae diseñada para usar la vía β-alanina y del 13% con una S. cerevisiae que utiliza la vía dependiente de la malonil-COA reductasa . Se determinó un mayor rendimiento de carbono de 3 HP por sustrato a través de la vía de la β-alanina en estudios con Escherichia coli que indicaron un valor del 42% en cultivos de glucosa en lotes alimentados .
Fig. 4
Comparación de parámetros de cultivo bajo diferentes limitaciones determinadas en quimiostatos de pequeña escala y reactores de sobremesa de 1 L en modo de alimentación por lotes. a rendimiento de carbono de 3 HP (%C-mol) y rendimiento de b de 3 HP en biomasa (g gCDW-1). Los errores corresponden a desviaciones estándar derivadas de cultivos triplicados
Para condiciones limitantes de P, sin embargo, nuestro estudio reveló un aumento considerable en el rendimiento de carbono de 3 HP de 25,6% (Fig. 4a). Los rendimientos de 3 HP observados en biomasa fueron de 0,38 y 0,65 g de gCDW – 1 para condiciones limitantes de C y P, respectivamente. Para la limitación de C, esto confirmó los valores observados en la configuración del quimiostato (Fig. 4b). Debido al aumento en el rendimiento de 3 HP por sustrato en condiciones limitantes de P y la ligera disminución en el rendimiento de biomasa por sustrato al mismo tiempo, el rendimiento de 3 HP por biomasa aumentó significativamente en comparación con el experimento de quimiostatos. A diferencia de los cultivos realizados en quimiostatos, no se detectó acumulación significativa de etanol o glicerol en cultivos alimentados por lotes. Un balance de carbono cerrado para las condiciones limitantes de C y P indicó que no se habían formado cantidades pertinentes de otros (subproductos)productos (cuadro 3). Esta diferencia en el espectro de subproductos podría explicar el aumento en el rendimiento de 3 HP por glucosa.
En comparación con los cultivos realizados en quimiostatos, las tasas específicas de absorción de glucosa determinadas en cultivos de lotes alimentados muestran valores considerablemente más altos de 0,75 y 0,77 mmol gCDW-1 h−1 para condiciones limitantes de C y P, respectivamente. Como se mencionó anteriormente, el punto de ajuste para el factor exponencial del perfil de alimentación se estableció en 0.05 h−1 y varió hasta 0.059 h−1, lo que resultó en una tasa de crecimiento específico 30-45% más alta en cultivos por lotes alimentados en comparación con los experimentos de quimiostatos con 0.04 h-1 (Tabla 3), causando un aumento de las tasas específicas de captación de glucosa. Sin embargo, para fermentaciones limitadas a C, el rendimiento de biomasa en glucosa fue comparable entre el cultivo de quimiostatos y el de lotes alimentados. De manera similar, las tasas de producción específicas de 3 HP fueron elevadas en cultivos por lotes alimentados, con 0,24 mmol de gCDW-1 h-1 y 0,38 mmol de gCDW−1 h−1, respectivamente. Los rendimientos de 3 HP por sustrato fueron comparables entre el cultivo de quimiostatos y el de lotes alimentados, lo que sugiere que los rendimientos son un parámetro de cultivo robusto que es resistente a las perturbaciones inducidas por fluctuaciones más pequeñas en la configuración del cultivo. No se detectó fosfato residual, ni glucosa, en las muestras tomadas de los diferentes reactores para condiciones limitantes de P, lo que confirma que los cultivos están limitados con respecto a la limitación correspondiente. Del mismo modo, para la limitación de C no se detectó glucosa residual en las muestras.
Las mejoras aquí presentadas de un sistema quimiostático a pequeña escala, desarrollado previamente por Klein et al. , que comprende, entre otras cosas, un aumento del número de reactores a 24 y una reducción del volumen de cultivo a 6,5 ml. Este mayor grado de paralelización hace que el sistema sea una herramienta de cribado de alto rendimiento adecuada, ya que varios sistemas de quimiostatos de pequeña escala comparables operan con volúmenes de trabajo más altos y con un número menor de reactores: El sistema de biorreactor continuo sacudido en paralelo (CosBios) utiliza seis recipientes de cultivo paralelos, respectivamente, con un volumen de cultivo de 20-25 ml . El sistema de biorreactor de tanque de agitación de un solo uso modificado para cultivos continuos (Biorreactor, 2mag AG) funciona con ocho reactores paralelos a un volumen de trabajo de 10 mL . El sistema de mini-quimiostato (MC) desarrollado por Bergenholm et al. consta de 16 reactores paralelos y requiere un volumen de trabajo de 40 ml. Por lo tanto, nuestro sistema es muy aplicable para la detección sencilla y rentable del rendimiento microbiano en modo continuo. Sin embargo, podría ser pertinente en mayor medida para el sistema mediante el seguimiento de parámetros adicionales, como los gases de escape, para permitir un análisis detallado de la distribución del carbono, que es de particular importancia para las caracterizaciones fisiológicas de deformación.
Como nuestro estudio sugirió que la limitación de N y P es favorable para la formación de 3 HP en S. cerevisiae, los experimentos futuros podrían implicar un mayor cribado de las tasas de crecimiento con la limitación de N y P aplicada para encontrar la producción óptima de 3 HP con cada limitación respectiva. Nuestro estudio mostró además la transferibilidad de los parámetros fisiológicos de los quimiostatos a los cultivos por lotes alimentados. Esto solo se considera factible si no se acumulan compuestos tóxicos o inhibidores durante el cultivo de los lotes alimentados, ya que los parámetros fisiológicos cambiarían con el tiempo y se desviarían de los parámetros adquiridos en los quimiostáticos. Como enfoque general, los experimentos con quimiostatos pueden servir como una herramienta para investigar la influencia de un compuesto potencialmente tóxico o inhibidor al agregar la sustancia al alimento en sí. Dado que todos los demás parámetros son constantes, la influencia del compuesto y su concentración pueden evaluarse y evaluarse directamente. En nuestro estudio, no se detectó formación de subproductos en cultivos de lotes alimentados y se indujo una respuesta al estrés presumiblemente similar debido a ácidos débiles, lo que permitió que las condiciones fisiológicas en quimiostáticos a pequeña escala se parecieran a las de los lotes alimentados.
En resumen, este estudio mostró que el concepto de condiciones limitantes de C y P para la producción de 3 HP se investigó en un novedoso sistema de cultivo de quimiostatos paralelizados y se pudo transferir con éxito a biorreactores de sobremesa de 1 L que operan en modo de alimentación por lotes. Por lo tanto, los parámetros fisiológicos adquiridos en los quimiostatos se pueden utilizar para el diseño y la evaluación del rendimiento de cultivos por lotes alimentados utilizando parámetros basados en el rendimiento para la configuración del proceso.