
Proteínas de adhesión celular
La adhesión celular-celular es un proceso selectivo, de tal manera que las células se adhieren solo a otras células de tipos específicos. Esta selectividad se demostró por primera vez en estudios clásicos del desarrollo embrionario, que mostraron que las células de un tejido (por ejemplo, el hígado) se adhieren específicamente a las células del mismo tejido en lugar de a las células de un tejido diferente (por ejemplo, el cerebro). Dicha adhesión selectiva celular-celular está mediada por proteínas transmembranas llamadas moléculas de adhesión celular, que se pueden dividir en cuatro grupos principales: las selectinas, las integrinas, la superfamilia de inmunoglobulinas (Ig) (así llamadas porque contienen dominios estructurales similares a las inmunoglobulinas) y las cadherinas (Tabla 12.3). La adhesión celular mediada por las selectinas, integrinas y cadherinas requiere Ca2 + o Mg2+, por lo que muchas interacciones adhesivas entre las células dependen de Ca2+ o Mg2+.
Cuadro 12.3 Moléculas de Adhesión Celular
.
Las selectinas median interacciones transitorias entre leucocitos y células endoteliales o plaquetas sanguíneas. Hay tres miembros de la familia de la selectina: la L-selectina, que se expresa en los leucocitos; la E-selectina, que se expresa en las células endoteliales; y la P-selectina, que se expresa en las plaquetas. Como se discutió anteriormente en este capítulo, las selectinas reconocen los carbohidratos de la superficie celular (ver Figura 12.14). Una de sus funciones críticas es iniciar las interacciones entre los leucocitos y las células endoteliales durante la migración de los leucocitos de la circulación a los sitios de inflamación de los tejidos (Figura 12.62). Las selectinas median la adhesión inicial de los leucocitos a las células endoteliales. Esto es seguido por la formación de adherencias más estables, en las que las integrinas en la superficie de los leucocitos se unen a moléculas de adhesión intercelular (iCams), que son miembros de la superfamilia Ig expresada en la superficie de las células endoteliales. Los leucocitos firmemente adheridos pueden penetrar las paredes de los capilares y entrar en el tejido subyacente al migrar entre las células endoteliales.

Figura 12.62
Adhesión entre leucocitos y células endoteliales. Los leucocitos abandonan la circulación en los sitios de inflamación de los tejidos al interactuar con las células endoteliales de las paredes capilares. El primer paso en esta interacción es la unión de selectinas leucocitarias (más…)
La unión de iCams a integrinas es un ejemplo de interacción heterofílica, en la que una molécula de adhesión en la superficie de una célula (por ejemplo, una ICAM) reconoce una molécula diferente en la superficie de otra célula (por ejemplo, una integrina). Otros miembros de la superfamilia Ig median interacciones homofílicas, en las que una molécula de adhesión en la superficie de una célula se une a la misma molécula en la superficie de otra célula. Tal unión homofílica conduce a la adhesión selectiva entre células del mismo tipo. Por ejemplo, las moléculas de adhesión de células nerviosas (N-CAMs) son miembros de la superfamilia Ig expresada en las células nerviosas, y la unión homofílica entre N-CAMs contribuye a la formación de asociaciones selectivas entre las células nerviosas durante el desarrollo. Hay más de 100 miembros de la superfamilia Ig, que median una variedad de interacciones célula-célula.
El cuarto grupo de moléculas de adhesión celular, las cadherinas, también muestran especificidades de unión homofílica. No solo participan en la adhesión selectiva entre células embrionarias, sino que también son los principales responsables de la formación de uniones estables entre células en los tejidos. Por ejemplo, la E-cadherina se expresa en las células epiteliales, por lo que las interacciones homofílicas entre las E-cadherinas conducen a la adhesión selectiva de las células epiteliales entre sí. Es de destacar que la pérdida de E-cadherina puede conducir al desarrollo de cánceres que surgen de las células epiteliales, lo que ilustra la importancia de las interacciones célula-célula en el control del comportamiento celular. Diferentes miembros de la familia de cadherinas, como la N-cadherina (cadherina neuronal) y la P-cadherina (cadherina placentaria), median la adhesión selectiva de otros tipos celulares.
Se han identificado unas veinte cadherinas clásicas diferentes, como la E-cadherina. Además, una subfamilia distinta de cadherinas, llamadas protocadherinas, se expresan en el sistema nervioso central, donde parecen desempeñar un papel en la adhesión entre las neuronas en las sinapsis. Curiosamente, diferentes neuronas parecen expresar diferentes protocadherinas, lo que sugiere que las protocadherinas pueden desempeñar un papel en el establecimiento de conexiones específicas entre neuronas. Alrededor de 50 genes de protocadherina humana han sido identificados y se ha demostrado que están organizados en tres grupos de genes. Cada grupo contiene múltiples exones que codifican los dominios extracelulares y de protocadherina transmembrana N-terminales, pero solo un solo conjunto de tres exones que codifican el dominio citoplasmático C-terminal (Figura 12.63). Por lo tanto, los grupos de genes de protocadherina parecen consistir en una región variable, que codifica múltiples dominios extracelulares y transmembranarios, vinculados a una región constante que codifica un único dominio citoplasmático. Esta organización de genes de protocadherina se asemeja notablemente a la de la inmunoglobulina y los genes receptores de células T (véanse las Figuras 5.42 y 5.43), en los que múltiples exones de región variable se unen a un solo exón de región constante. En los genes receptores de inmunoglobulina y células T, esto ocurre como resultado de reordenamientos del ADN que generan diversidad en el sistema inmunitario. Queda por determinar si las regiones variables y constantes de las protocadherinas están unidas a nivel de ADN o ARN (por ejemplo, mediante empalmes alternativos) y en qué medida los reordenamientos de los genes de la protocadherina podrían contribuir al establecimiento de conexiones sinápticas específicas en el cerebro.
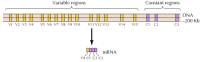
Figura 12.63
Organización de grupos de genes de protocadherina. Los genes de la protocadherina humana están organizados en tres grupos. En el cluster ilustrado, 15 regiones variables diferentes que codifican dominios extracelulares y transmembranosos están vinculadas a una sola región constante, (más…)
En contraste con las uniones estables de matriz celular discutidas en la sección anterior, las interacciones célula-célula mediadas por las selectinas, integrinas y miembros de la superfamilia Ig son adherencias transitorias en las que los citoesqueletos de las células adyacentes no están vinculados entre sí. Las uniones de adhesión estables que involucran los citoesqueletos de las células adyacentes están mediadas por las cadherinas. Como se discutió en el Capítulo 11, estas uniones entre células son de dos tipos: uniones adherentes y desmosomas, en las que las cadherinas o proteínas relacionadas (desmogleínas y desmocolinas) están unidas a haces de actina y filamentos intermedios, respectivamente (Figura 12.64). El papel de las cadherinas en la unión de los citoesqueletos de las células adyacentes es análogo al de las integrinas en la formación de uniones estables entre las células y la matriz extracelular.

Figura 12.64
Uniones célula-célula estables mediadas por las cadherinas. Las interacciones homofílicas entre cadherinas median dos tipos de adherencias célula-célula estables. En las uniones adherentes, las cadherinas están unidas a haces de filamentos de actina a través de las cateninas (ver Figura (más…)